Abstract
Human cannabinoid receptor CB2 is an integral membrane protein that belongs to a large superfamily of seven transmembrane domain G protein-coupled receptors (GPCR). CB2 is a part of the endocannabinoid system that regulates immune response, inflammation and pain sensitivity. The rational development of novel specific pharmaceuticals targeting this receptor relies on high resolution studies of the structure and mechanisms of activation of CB2 . Here we review the methodology for recombinant expression and purification of CB2 , suitable for preparation of multi-milligram quantities of functionally active receptor. CB2 is expressed in a functional form in E. coli, solubilized in detergent micelles, and purified by tandem affinity chromatography. The recombinant CB2 is amenable to functional and structural studies by nuclear magnetic resonance spectroscopy and a wide range of biochemical and biophysical techniques.
Introduction
Heptahelical G protein-coupled receptors (GPCR) are integral membrane proteins involved in a wide array of cell signaling pathways. The cannabinoid receptor CB2 that primarily expresses in cells of immune origin is an attractive target for the development of drugs for management of inflammation, immunological disorders and pain.1-3 Structural studies can provide critical contribution to the rational design of novel specific drugs targeting this receptor.
The progress in structural studies of GPCR has been relatively slow, primarily due to the difficulties in obtaining large quantities of sufficiently pure, homogenous and functional receptors as well as conformational flexibility of GPCR that complicates preparation of well-diffracting crystals for X-ray crystallography. Recent improvements in expression techniques, methods of stabilization and crystal preparation resulted in over 30 high resolution structures of GPCR solved during the past few years, advancing our understanding of cell signaling.4-6 In addition to X-ray crystallography, a large array of biophysical methods have been employed to study the structure and function of these receptors. In particular, applications of nuclear magnetic resonance (NMR) gain increasing attention since they allow studies of structurally unperturbed GPCR in native-like environments such as lipid bilayers.
With the exception of rhodopsin, most GPCR are present in native tissues at relatively low levels, and recombinant expression in a heterologous host is the only practical way to obtain these proteins in quantities required for structural studies. The commonly used expression systems include baculovirus-infected insect cells, yeast, bacterial or mammalian cells as well as cell-free systems.7-12 However, the application of NMR to studies of GPCR require either site-specificor uniform labeling of these proteins with stable isotopes.12 This can be achieved via metabolic labeling, through co-translational incorporation of labeled amino acids into the protein. The methods of stable isotope-labeling of proteins via expression in E. coli have been well developed,13-15 while the use of insect- or mammalian cells for preparation of stable-isotope labeled proteins only recently became of practical interest, due to the high cost and complexity of the growth medium supplemented with labeled nutrients.16 The expression in yeast cells has also been used for production of several GPCR,17 but this host may not be suitable for preparation of some receptors because of the non-homogenous glycosylation or partial proteolysis of target proteins. Thus, the stable isotope labeling by expression in E. coli cells is currently the most cost-effective, practical alternative. Here we review the methodological approaches for expression and purification of CB2 . These methodst are suitable for preparation of milligram quantities of functional GPCR for studies by NMR and a broad array of other biophysical techniques.
Functional Expression in E. coli
The general outline of the experimental strategy is given in Figure 1. Protein is expressed in E. coli membranes, and the expression levels and functional activity assessed by Western blot, ligand binding and G protein-activation assays.18-20 The purification is performed upon solubilization of the receptor in detergent micelles followed by the two rounds of affinity chromatography. The purified receptor can be studied either when solubilized in detergent micelles or after reconstitution into lipid bilayers in a form of proteoliposomes or nanodiscs.
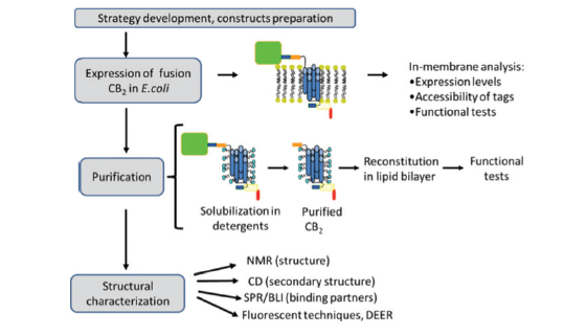 Figure 1. Experimental strategy for preparation of functional CB2
Figure 1. Experimental strategy for preparation of functional CB2Multiple factors such as the choice of the bacterial host and expression vector, strength of the promoter, composition of culture media, temperature and length of the induction determine both the total yield as well as the fraction of correctly folded functional receptor. Several E. coli strains including BL21 (DE3), DH5a, KRX, Rosetta Gami, C41 and C43 were compared for their effectiveness in production of fusion CB2 ; strain BL21 (DE3) provided the highest yield of the functional receptor.21, 22
The expression partners fused to the target protein at the N-and C-termini can also dramatically affect the expression level and activity of the recombinant GPCR. A variety of plasmid constructs containing different combinations of expression- and solubility tags were tested for their efficiency in production of functional CB2 . Figure 2 depicts selected expression constructs currently utilized in our laboratory.
We established that for proper folding and functionality of the recombinant CB2 in E. coli, an appropriate N-terminal fusion partner is essential.19 For example, the N-terminal Haloalkane dehalogenase (Halotag) significantly increased the levels of the fusion CB2 protein. However, the accumulated receptor was not functional.24 The expression of CB2 without the N-terminal fusion partner resulted in only trace quantities of the recombinant protein.19 On the other hand, the N-terminal maltose-binding protein (MBP) was highly beneficial for the high-level functional expression of this receptor.21 The MBP is localized in the periplasm of E. coli; upon leaving the ribosome the nascent peptide is transported across the cytoplasmic membrane by the Sec translocon system.25 This may facilitate the insertion of the downstream hydrophobic part of the fusion protein into cytoplasmic membranes. Importantly, the full length sequence of MBP is required for the maximal beneficial effect.19
The topology of the receptor in cytoplasmic membranes was determined by measuring the in vivo biotinylation of the fusion protein containing a biotin-recognition sequence placed in various positions relative to CB2 . 26 When expressed as a fusion with MBP, the N-terminus of CB2 is exposed to the periplasmic space while its C-terminal part is localized to the cytoplasm of E. coli. While the MBP is believed to promote the insertion of the fusion protein into cytoplasmic membranes in an N-terminus-out orientation, the oxidative redox environment of the periplasm may facilitate the formation of the essential disulfide bond in extracellular loop,2 thereby stabilizing CB2 . The expression of the recombinant protein can be further increased by 30-40% by fusing the E. coli thioredoxin (Trx) to the C-terminus of CB2 . Both MBP and Trx expression partners can be selectively removed if desired by cleaving the fusion with the specific TEV protease at recognition sequences flanking the receptor.
To enable the purification of CB2 by tandem affinity chromatography, two small affinity tags can be added at the opposing ends of the recombinant protein. Examples of several such constructs are shown in Figure 2 and include His- and Strep-tag pair, His-tag/Bio-tag and Rho-tag/His-tag pairs, to name a few.
 Figure 2. Schematic representation of constructs for expression of CB2 fusion protein in E. coli. (adapted from 12,19,23)
Figure 2. Schematic representation of constructs for expression of CB2 fusion protein in E. coli. (adapted from 12,19,23)The expression of CB2 is performed from the low copy number vector based on the pMal-p2 backbone, from the weak lac promoter. The rather low rates of transcription allow better alignment of the protein synthesis with the rate of folding of the nascent polypeptide and its insertion into cytoplasmic membranes. For the same reason, lowering of the cultivation temperature from 37 °C to 20 °C has a clear beneficial effect for the functional expression of CB2 , improving the yield of ligand binding-competent receptor from less than 20-30% to close to 100%.22 Please note though that further decrease of induction temperature to below 20 °C leads to significant drop in the protein expression levels and therefore is not practical.22
For stable-isotope labeling of CB2 a special protocol involving the expression in a minimal salt medium (MSM) supplemented with labeled nutrients was developed.22,27 In order to compensate for the decrease of expression levels of the target protein in MSM, the cultivation of the E. coli cells in a fermenter was performed. Either glucose and ammonium salts or a mixture of amino acids can be used as sources of carbon and nitrogen. Under the controlled conditions of aeration, pH and nutrient supply the yield of the biomass in the fermenter can be increased 10-fold or higher, compared to the expression in rich 2xYT medium in shake-flasks. As much as 2-3 mg of the recombinant receptor can be obtained in 1L of fermentation culture.
Purification of the Recombinant CB2
Purification of CB2 is performed following its solubilization in detergent micelles. By screening solubilization conditions with ~ 40 different detergents, a mixture of the nonionic dodecyl maltoside (DDM) and zwitterionic CHAPS was identified as the most suitable for efficient extraction of the CB2 from membranes (Table 1). Further supplementation with cholesteryl hemisuccinate (CHS) and a high affinity ligand results in significant stabilization of the receptor in detergent micelles.18
Table 1. Detergents for solubilization of recombinant CB2.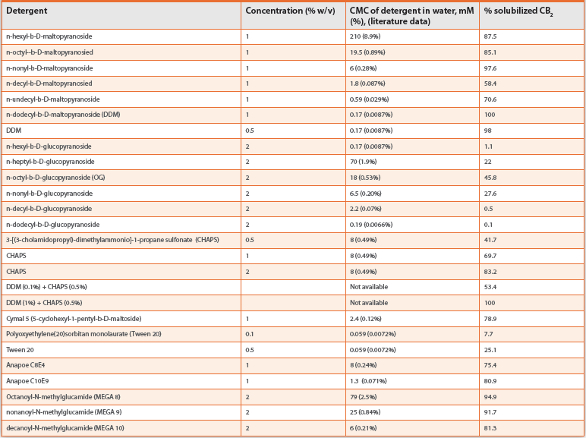
Even when cultivated in a rich 2xYT medium, the E. coli cells accumulate CB2 at moderate levels (~0.1% of total cellular protein) (21). The cultivation in a defined MSM medium results in even lower CB2 content. Thus, a very efficient and selective chromatographic procedure is required to achieve the yield and purity of the target protein required for high resolution structural methods. Such chromatographic techniques as ion-exchange, size-exclusion or hydrophobic chromatography were not suitable, since the large detergent micelles prevented proper interaction of the receptor with the resin. On the other hand, high level purification and recovery of CB2 was achieved by tandem affinity chromatography taking advantage of small affinity tags placed at opposing ends of the receptor.19
A typical purification protocol for fusion constructs of CB2 containing C-terminal decahistidine tags begins with the IMAC chromatography. The binding of a shorter histidine tag (i.e. hexahistidine) to the Ni-NTA resin in the presence of detergent micelles was shown to be rather weak,21 while the longer decahistidine tag makes this purification step much more efficient and typically results in a recovery of ~80% or more of the fusion CB2 . The fusion partners such as MBP can be removed after the elution from the resin, by cleaving the fusion protein with specific TEV protease at sites flanking the receptor.
The CB2 released from the fusion is then re-captured via the N-terminal affinity tag (such as StrepTag), to separate it from TEV protease, cleavage products and other impurities. The capture of CB2 on the affinity resin is performed in the presence of detergents DDM, CHAPS and CHS as well as salts and glycerol that stabilize the receptor. Since the affinity of a single Strep-tag (WSHPQFEK ) to the resin in the presence of detergent micelles is weak, a double repeat of this tag (twin-Strep-tag) is used to ensure a more efficient capture.19 The beneficial avidity effect was reported also for purification of soluble proteins on a StrepTactin resin.28 This double tag typically improves the recovery of the receptor to greater than 70-75%.
The expression of CB2 in a defined minimal salt medium for stable isotope-labeling decreases the titer of the recombinant protein. To maximize the recovery of the target protein, the use of high affinity purification resin is highly desirable. The recently released StrepTactin XT resin satisfies this requirement, since it has an estimated affinity of interaction with the twin-Strep-tag in the low-nanomolar range (Figure 3).29 The retention of the captured protein on a resin is very strong, which allows stringent washing conditions. The elution of the captured CB2 is performed at mild conditions, upon addition of 50 mM biotin to the running buffer. This purification protocol allows recovery of up to 80% of the recombinant CB2 originally present in the cell extracts, with high degree of purity (over 90%).
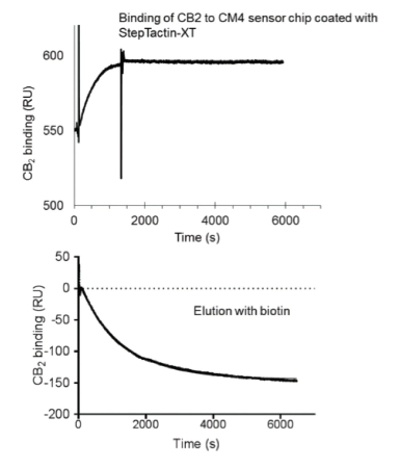 Figure 3. Surface plasmon resonance studies of binding of twin- Strep-tagged CB2 to StrepTactin XT.
Figure 3. Surface plasmon resonance studies of binding of twin- Strep-tagged CB2 to StrepTactin XT.The recovered protein is highly pure and functionally active, as supported by the results of Ligand binding and G protein-activation experiments (Figure 4).
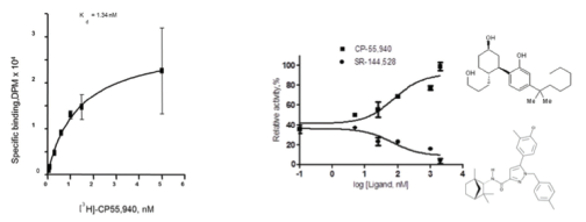 Figure 4. A, Ligand binding on CB2 reconstituted into liposomes. B, activation of G protein on CB2 reconstituted into liposomes in the presence of agonist CP-55,940 (upper curve) or inverse agonist SR-144528 (lower curve).
Figure 4. A, Ligand binding on CB2 reconstituted into liposomes. B, activation of G protein on CB2 reconstituted into liposomes in the presence of agonist CP-55,940 (upper curve) or inverse agonist SR-144528 (lower curve).To summarize, the protocols developed for recombinant production of CB2 in E. coli cells and purification using tandem affinity chromatography yield the fully functional protein at high purity. Optimization of the expression plasmid, cultivation conditions and other parameters resulted in the recovery of the recombinant CB2 as high as 2-3 mg of protein from 1 liter of culture. The technology of preparation of the recombinant CB2 can be adapted to expression and purification of other GPCR and recombinant receptors. The developed methods for expression, stable isotope labeling, purification, stabilization and liposome reconstitution open a path to meaningful NMR spectroscopic studies on the CB2 receptor.
Acknowledgements
This work was supported by the Intramural Research Program of the NIAAA, NIH.
References
- Graham, E. S., Ashton, J. C., and Glass, M. (2009) Cannabinoid receptors: A brief history and “what’s hot”, Frontiers in Bioscience-Landmark 14, 944-957.
- Munro, S., Thomas, K. L., and Abushaar, M. (1993) Molecular characterization of a peripheral receptor for cannabinoids, Nature 365, 61-65.
- Pacher, P., and Mechoulam, R. (2011) Is lipid signaling through cannabinoid 2 receptors part of a protective system?, Prog Lipid Res 50, 193-211.
- Lefkowitz, R. J. (2013) A brief history of G-protein coupled receptors (Nobel Lecture), Angewandte Chemie 52, 6366-6378.
- Thal, D. M., Sun, B., Feng, D., Nawaratne, V., Leach, K., Felder, C. C., Bures, M. G., Evans, D. A., Weis, W. I., Bachhawat, P., Kobilka, T. S., Sexton, P. M., Kobilka, B. K., and Christopoulos, A. (2016) Crystal structures of the M1 and M4 muscarinic acetylcholine receptors, Nature 531, 335-340.
- Vaidehi, N., Grisshammer, R., and Tate, C. G. (2016) How Can Mutations Thermostabilize G-Protein-Coupled Receptors?, Trends in pharmacological sciences 37, 37-46.
- Granier, S., Manglik, A., Kruse, A. C., Kobilka, T. S., Thian, F. S., Weis, W. I., and Kobilka, B. K. (2012) Structure of the delta-opioid receptor bound to naltrindole, Nature 485, 400-404.
- Jaakola, V.-P., Griffith, M. T., Hanson, M. A., Cherezov, V., Chien, E. Y. T., Lane, J. R., Ijzerman, A. P., and Stevens, R. C. (2008) The 2.6 Angstrom crystal structure of a human A(2A) adenosine receptor bound to an antagonist, Science 322, 1211-1217.
- Mancia, F., and Hendrickson, W. A. (2007) Expression of recombinant G-protein coupled receptors for structural biology, Mol bioSys 3, 723-734.
- Rasmussen, S. G. F., Choi, H.-J., Fung, J. J., Pardon, E., Casarosa, P., Chae, P. S., DeVree, B. T., Rosenbaum, D. M., Thian, F. S., Kobilka, T. S., Schnapp, A., Konetzki, I., Sunahara, R. K., Gellman, S. H., Pautsch, A., Steyaert, J., Weis, W. I., and Kobilka, B. K. (2011) Structure of a nanobody-stabilized active state of the beta(2) adrenoceptor, Nature 469, 175-180.
- Rosenbaum, D. M., Zhang, C., Lyons, J. A., Holl, R., Aragao, D., Arlow, D. H., Rasmussen, S. G. F., Choi, H.-J., DeVree, B. T., Sunahara, R. K., Chae, P. S., Gellman, S. H., Dror, R. O., Shaw, D. E., Weis, W. I., Caffrey, M., Gmeiner, P., and Kobilka, B. K. (2011) Structure and function of an irreversible agonist-beta(2) adrenoceptor complex, Nature 469, 236-240.
- Yeliseev, A. A., and Vukoti, K. (2011) Expression of G protein-coupled receptors, In Production of membrane proteins (Robinson, A. S., Ed.), pp 219-248, Wiley-VCH, Weinheinm, Germany.
- Ross, A., Kessler, W., Krumme, D., Menge, U., Wissing, J., van den Heuvel, J., and Flohe, L. (2004) Optimised fermentation strategy for 13C/15N recombinant protein labelling in Escherichia coli for NMR-structure analysis, Journal of biotechnology 108, 31-39.
- Cai, M., Huang, Y., Sakaguchi, K., Clore, G. M., Gronenborn, A. M., and Craigie, R. (1998) An efficient and cost-effective isotope labeling protocol for proteins expressed in Escherichia coli, Journal of biomolecular NMR 11, 97-102.
- Marley, J., Lu, M., and Bracken, C. (2001) A method for efficient isotopic labeling of recombinant proteins, Journal of biomolecular NMR 20, 71-75.
- Casagrande, F., Maier, K., Kiefer, H., Opella, S. J., and Park, S. H. (2011) Expression and purification of G-protein-coupled receptors for nuclear magnetic resonance structural studies, In Production of membrane proteins (Robinson, A. S., Ed.), pp 297-316, WileyVCH, Weinheim, Germany.
- Britton, Z., Young, C., Can, O., McNeely, P., Naranjo, A., and Robinson, A. S. (2011) Membrane protein expression in Saccharomyces cerevisiae, In Production of membrane proteins (Robinson, A. S., Ed.), pp 37-74, Wiley-VCH, Weinheim, Germany.
- Vukoti, K., Kimura, T., Macke, L., Gawrisch, K., and Yeliseev, A. (2012) Stabilization of functional recombinant cannabinoid receptor CB2 in detergent micelles and lipid bilayers, PLOS One 7, e46290.
- Yeliseev, A., Zoubak, L., and Gawrisch, K. (2007) Use of dual affinity tags for expression and purification of functional peripheral cannabinoid receptor, Prot Express Purif 53, 153-163.
- Kimura, T., Yeliseev, A. A., Vukoti, K., Rhodes, S. D., Cheng, K., Rice, K. C., and Gawrisch, K. (2012) Recombinant cannabinoid type 2 receptor in liposome model activates g protein in response to anionic lipid constituents, The Journal of biological chemistry 287, 4076-4087.
- Yeliseev, A. A., Wong, K. K., Soubias, O., and Gawrisch, K. (2005) Expression of human peripheral cannabinoid receptor for structural studies, Prot Sci 14, 2638-2653.
- Berger, C., Ho, J. T. C., Kimura, T., Hess, S., Gawrisch, K., and Yeliseev, A. (2010) Preparation of stable isotope-labeled peripheral cannabinoid receptor CB2 by bacterial fermentation, Prot Express Purif 70, 236-247.
- Locatelli-Hoops, S., Gorshkova, I., Gawrisch, K., and Yeliseev, A. A. (2013) Expression, surface immobilization, and characterization of functional recombinant cannabinoid receptor CB2 , BBA-Proteins and Proteomics 1834, 2045-2056.
- Locatelli-Hoops, S., Sheen, F. C., Zoubak, L., Gawrisch, K., and Yeliseev, A. A. (2013) Application of HaloTag technology to expression and purification of cannabinoid receptor CB2 , Prot Express Purif 89, 62-72.
- Calandra, B., Tucker, J., Shire, D., and Grisshammer, R. (1997) Expression in Escherichia coli and characterisation of the human central CB1 and peripheral CB2 cannabinoid receptors, Biotechnol Lett 19, 425-428.
- Krepkiy, D., Wong, K., Gawrisch, K., and Yeliseev, A. (2006) Bacterial expression of functional, biotinylated peripheral cannabinoid receptor CB2 , Prot Express Purif 49, 60-70.
- Kimura, T., Vukoti, K., Lynch, D. L., Hurst, D. P., Grossfield, A., Pitman, M. C., Reggio, P. H., Yeliseev, A. A., and Gawrisch, K. (2014) Global fold of human cannabinoid type 2 receptor probed by solid-state 13C-, 15N-MAS NMR and molecular dynamics simulations, Proteins 82, 452-465.
- Schmidt, T. G. M., Batz, L., Bonet, L., Carl, U., Holzapfel, G., Kiem, K., Matulewicz, K., Niermeier, D., Schuchardt, I., and Stanar, K. (2013) Development of the Twin-Strep-tag (R) and its application for purification of recombinant proteins from cell culture supernatants, Protein Expres Purif 92, 54-61.
- Yeliseev, A., Zoubak, L., and Schmidt, T. G. (2016) Application of Strep-Tactin XT for affinity purification of Twin-Strep-tagged CB2 , a G protein-coupled cannabinoid receptor, Protein Expr Purif., in press.
Author Biography
Alexei Yeliseev is a Staff Scientist and the head of the protein chemistry group at the National Institute on Alcoholism and Alcohol Abuse, National Institute of Health. His research focuses on developing technologies for expression, purification, and functional and structural characterization of G protein-coupled receptors.