Introduction
Glycans, also known as oligosaccharides, are complex, highly branched carbohydrate structures composed of monosaccharide sugars such as fucose, galactose, mannose, N-acetylglucosamine and sialic acid (N-acetylneuraminic acid). They are typically found either attached to proteins (glycoproteins) or lipids (glycolipids). Glycosylation, the process of adding sugar units to a protein, is a common post-translational modification (PTM) in eukaryotic cells.1 There are two main types of glycans: N-linked glycans, which are bound to the nitrogen on the amino acids asparagine or arginine, and O-linked glycans, which are bound to the hydroxyl oxygen on the amino acids serine or threonine.
Although there is still much to learn about glycans, it is widely believed that they may affect a protein’s functionality and critical properties such as binding, stability, folding, immunogenicity and turnover. For example, the Zika virus has been covered widely in the news over the last several months. Purdue University researchers recently reported that they have determined the structure of the Zika virus. The team found that the vicinity of the glycosylation site in the Zika virus differs from other similar viruses.2 Different glycosylation of the Zika virus could be the reason why it can attach to human cell receptors in a different manner and be able to penetrate blood-brain and placental barriers. This is a significant finding, and understanding this mechanism may help in developing a vaccine.
Importance of Glycosylation
Glycosylation is a Critical Quality Attribute (CQA) that must be presented to ensure safety and potency of mAbs and other biopharmaceutical products before regulatory approval.3 The presence of glycosylation affects product stability,4 immunogenicity (ADCC, antibody-dependent cell-mediated cytotoxicity, and CDC, complement dependent cytotoxicity),5,6 serum clearance,7 pharmacokinetics8 and may affect anti-inflammatory activity.9,10 The structure-function relationship due to the complex glycosylation patterns is still not understood; therefore, it is imperative to obtain the full glycan profile of a glycoprotein during discovery, clinical and manufacturing phases.
Methods to Determine Glycan Profiles
Numerous approaches are employed to characterize glycans, and along with these there are also many analytical challenges and considerations. However, the classical approach to glycan analysis is to remove the N-linked glycans from the protein prior to analysis using an enzyme such as Peptide N-Glycosidase F. More exotic enzymes are required for removing O-linked glycans from proteins.11 Unfortunately there is no general O-glycosidase that removes all O-glycans completely from a sample in their intact forms. Therefore, if intact O-glycan profiling is needed, a chemical method (like a mild hydrazinolysis) is required.12 Once released, the glycans, which have no UV chromophores, cannot be detected by common quality control lab detectors like UV detectors without derivatization. To maximize sensitivity, the glycans are usually derivatized with a fluorophore. The entire glycan preparation can be tedious, labor intensive, and technique-dependent. Recently developed commercial glycan kits and novel labeling reagents have helped to make the preparation process more efficient and less time consuming. Separation of various glycan components is also challenging due to their hydrophilic nature and similar composition. Eurofins Lancaster Laboratories recently participated in a NIST Interlaboratory Study on Glycosylation Analysis to explore results obtained by a wide range of analytical approaches. Hydrophilic Interaction Liquid Chromatography (HILIC) utilizing shallow gradients associated with long run times is often used to separate the numerous glycans as illustrated in Figure 1. A longer analysis using a shallow gradient provides better resolution, but at the expense of time.
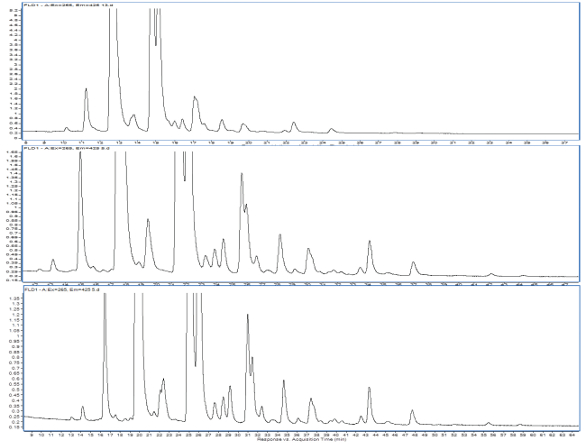 Figure 1. The shallowest gradient shown in the bottom results in better HILIC separation: The glycan profile of the NIST antibody (Rapifluor® labeling). Mobile phase of acetonitrile changed in composition from 75% to 60% over 20 (top frame), 50 (middle frame) and 75 (bottom frame) minutes.
Figure 1. The shallowest gradient shown in the bottom results in better HILIC separation: The glycan profile of the NIST antibody (Rapifluor® labeling). Mobile phase of acetonitrile changed in composition from 75% to 60% over 20 (top frame), 50 (middle frame) and 75 (bottom frame) minutes.According to the NIST Interlaboratory Study on Glycosylation Analysis, the two most popular methods are analyzing released glycans (~74%) and glycopeptides (~20%) carrying the glycans. For fluorescence derivatization approaches, the released glycans are equally derivatized using 2-aminobenzamide (2-AB) tag or glycosylamine labeling techniques (RapiFluor® or InstantPC), preceding all other derivatization techniques (APTS, 2-AA, Procainamide). The 2-AB label provides extra hydrophobicity to glycan molecules; therefore, they are suitable for both reversed-phase and hydrophilic interaction liquid chromatography. These two separations yield orthogonal and complementary results that may be beneficial in a complex separation. Additionally, an extensive database for 2-AB-labeled glycans exists due to its popularity and long-term use in the analytical community, offering a valuable aid in glycan identification.13 When properly separated to avoid closely eluting species, the standardized elution order supports identification when mass spectrometric identification is not straightforward. Recently, novel labeling agents have gained popularity due to their fast labeling process (RapiFluor®, InstantPC), enhanced fluorescence sensitivity, and more desirable ionization properties for mass spectrometry (RapiFluor®, InstantPC, Procainamide). All these labels contain a tertiary amine that provides efficient electrospray ionization producing a strong MS response.
The majority of the therapeutic proteins are glycoproteins (monoclonal antibodies, blood proteins, growth factors, vaccines, fusion proteins).14 For example erythropoietin (EPO), a common biotherapeutic protein, is highly glycosylated and may contain up to 50% glycans by weight, having multiple sites of glycosylation (three N-linked glycan sites occupied with highly branched and sialylated glycans and one O-linked glycan).15 A monoclonal antibody (mAb) such as an IgG1 has a single site of attachment usually at the asparagine sequence specific to the NXS/T motif inside the CH2 part of Fc domain on the heavy chains and exhibits a much simpler glycan profile. For example the N-linked glycan profile of an IgG1 mAb reference standard (Waters Corp.) derivatized with RapiFluor® is presented in Figure 2.
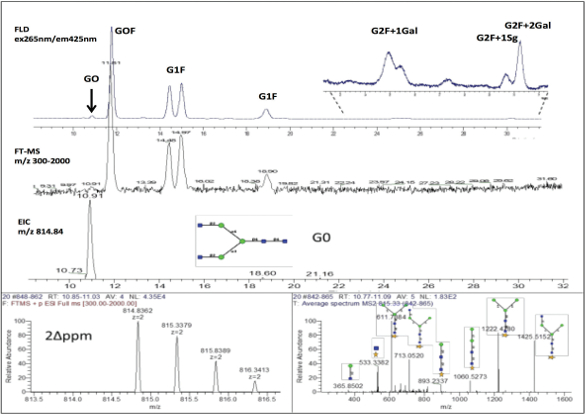 Figure 2. The glycan profile (fluorescence) of the IgG1 mAb reference standard labeled with RapiFluor® (upper frame), the corresponding TIC chromatogram (middle frame) and m/z 814.84 EIC for G0 (bottom frame) with example MS and MS2 spectra for G0 (bottom).
Figure 2. The glycan profile (fluorescence) of the IgG1 mAb reference standard labeled with RapiFluor® (upper frame), the corresponding TIC chromatogram (middle frame) and m/z 814.84 EIC for G0 (bottom frame) with example MS and MS2 spectra for G0 (bottom).Proteins with complex glycosylation profile can include fusion proteins, especially when a growth factor protein is attached to the Fc region of an antibody as the case with Enbrel,16 which has an approved biosimilar version on the market. An example glycan profile of a large fusion protein (>200 kDa) that is approximately 20% glycosylated is presented in Figure 3.
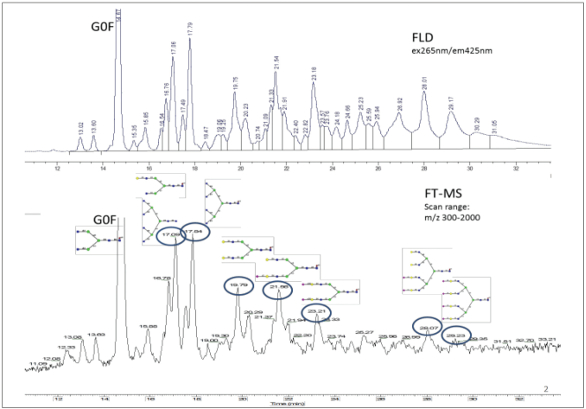 Figure 3. Fluorescence glycan profile of a heavily glycosylated 200 kDa fusion protein (top) and corresponding TIC chromatogram (bottom).
Figure 3. Fluorescence glycan profile of a heavily glycosylated 200 kDa fusion protein (top) and corresponding TIC chromatogram (bottom).Glycan Identification Using Mass Spectrometry
Mass spectrometry can be used to identify glycans in complex fluorescence chromatograms. The LC flow is either split between or in series with a high resolution mass spectrometer that provides accurate mass through the MS spectra or structural information through the MS2 spectra. The identification of the glycan peaks shown in Figure 2 and Figure 3 included determining the molecular mass for each significant fluorescence peak by analyzing the MS spectra generated by a Fourier Transform Mass Spectrometer (FTMS Orbitrap) and comparing the extracted ion chromatograms (EIC). Glycan identification was assigned based on historical record of the expected elution order and the mass accuracy (Δppm<10) between the observed mass and theoretical glycan mass as illustrated in Figure 2. In addition to the described workflow, tandem mass spectrometry (MS/MS) after collision induced dissociation (CID) can be used to help determine glycan structure and to differentiate between isobaric species.
Glycosylation can be evaluated at the protein or peptide level using high resolution mass spectrometry using a top down approach and/or a middle down approach.17 The intact molecule or its subunits can be analyzed to identify glycoforms and estimate their relative abundances. Figure 4 shows a deconvoluted mass spectrum of the FC/2 domain of adalimumab after a digestion with Ides enzyme. Both electrospray ionization (ESI) and MALDI-mass spectrometry can be used for analysis of glycans at the protein and peptide level. In top-down analyses, high resolution is required to analyze the many proteoforms and glycoforms of the same polypeptide chain. Electrospray mass spectrometry is most often used in-line with reverse-phase or HILIC separation along with MS-friendly mobile-phases or CE separation, whereas MALDI-MS is a stand-alone technique offering a quick screening tool for complex mixtures.
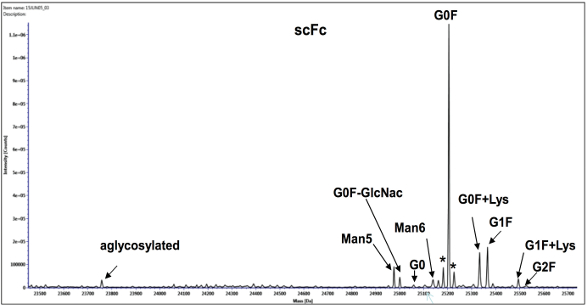 Figure 4. Deconvoluted mass spectrum showing the glycoforms from a single chain of the Fc (scFc) of region of the heavy chain of Adalimumab after reduction and digestion with Ides enzyme.
Figure 4. Deconvoluted mass spectrum showing the glycoforms from a single chain of the Fc (scFc) of region of the heavy chain of Adalimumab after reduction and digestion with Ides enzyme.A related analysis performed in our laboratory is sialic acid quantitation. Sialic acid is a generic name for a unique group of over 30 9-carbon monosaccharides that attach to the ends of glycans. N-Acetylneuraminic acid (NANA) and N-Glycolylneuraminic acid (NGNA) are the two common sialic acids found in humanized antibodies (99% usually is NANA). It is critical to measure sialic acid content as it can have significant therapeutic implications.9,10
Evaluation of glycosylation as a quality attribute requires the identification of glycan structures and their abundance as well as locating the sites of glycosylation on the protein.18 Bottom up approaches after protease digestion using liquid chromatography and tandem mass spectrometer (LC/MS/MS) allow for the analysis of glycopeptides to evaluate glycan structures and amino acid sequences at the site of glycosylation.19 Traditional collision induced dissociation (CID) allows identification of the N-glycosylation site based on comparison of LC/MS/MS data of the glycosylated and deglycosylated protein digests. The mass shift of 0.98 Da on specific Asn residues after PNGaseF digestion indicates hydrolysis of the bond between Asn and the GlcNac group of the N-glycan. The use of electron transmission dissociation (ETD) displays c and z ions from the glycopeptide backbone and provides spectral information for locating the glycan position in the protein. Nano flow chromatography and ionization can improve glycopeptide separation and increases sensitivity during their detection.20 When these techniques are used in conjunction with modern scanning mass spectrometers capable of multiple types of fragmentation, maximum information concerning the amino acid sequence, the site of glycosylation, and glycan structure can readily be obtained in a single analysis.
Conclusion
Glycan profiling is extremely complex and provides essential characterization information about therapeutic proteins and biosimilars. Because glycosylation is one of the most important post-translation modifications that occur during production, this quality attribute must be monitored experimentally during the lifetime of the biotherapeutic and cannot be predicted based on the gene sequence or protein chemistry.21 The complexity of glycans that can be added to a protein contributes to its heterogeneity, opening the unanswerable question as to the biological significance of the multitude of glycan peaks that can appear during profiling.22
Eurofins Lancaster Laboratories has significant experience analyzing glycans. Information from glycan analysis can be used at all stages of drug development to determine identities and quantities of the various monosaccharides/glycans present, type of glycosylation (N-linked and/or O-linked), extent to which glycosylation has occurred, consistency between batches for release during the manufacturing process, comparison of glycoproteins manufactured using different cell lines, and specifications for stability and release.
References
- Apweiler R, Hermjakob H, Sharon N. On the frequency of protein glycosylation, as deduced from analysis of the SWISS-PROT database. Biochim Biophys Acta 1999;1473(1):4-8.
- Shirohi, D, Chen Z., Sun L, Klose T, Pierson TC, Rossmann MG, Kuhn RJ. The 3.8 Å resolution cryo-EM structure of Zika virus. Science 2016;352(6284):467-70.
- Zhang P, Woen S, Wang T, et. al. Challenges of glycosylation analysis and control: an integrated approach to producing optimal and consistent therapeutic drugs. Drug Discov. Today 2016;21(5):740-65.
- Sola RJ, Griebenow K. Effects of Glycosylation on the Stability of Protein Pharmaceuticals. J Pharm Sci. 2009;98(4):1223–1245.
- Natsume A, Niwa R, Satoh M. Improving effector functions of antibodies for cancer treatment: Enhancing ADCC and CDC. Drug Des Devel Ther. 2009;21(3):7-16.
- Quast I, Keller CW, Maurer MA, et. al. Sialylation of IgG Fc domain impairs complementdependent cytotoxicity. J Clin Invest. 2015;125(11):4160-70.
- Goetze AM, Diana Liu YD, Zhang Z, et al. High-mannose glycans on the Fc region of therapeutic IgG antibodies increase serum clearance in humans. Glycobiology 2011;21(7):949-59.
- Higel F, Seidl A, Sörgel F, Friess W. N-glycosylation heterogeneity and the influence on structure, function and pharmacokinetics of monoclonal antibodies and Fc fusion proteins. European J. of Pharm. Biopharm. 2016;100:94-100.
- Samuelsson A, Towers TL and Ravetch JV. Anti-inflammatory activity of IVIG mediated through the inhibitory Fc receptor. Science 2001;291(5503):484–486
- Nagelkerke SQ, Dekkers G, Kustiawan I, et. al. Inhibition of FcγR-mediated phagocytosis by IVIg is independent of IgG-Fc sialylation and FcγRIIb in human macrophages. Blood 2014;124(25):3709-3718.
- Varki A, Cummings RD, Esko JD, et. al. Essentials of Glycobiology. 2nd edition. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press; 2009.
- Royle L, Mattu TS, Hart E, et. al. An analytical and structural database provides a strategy for sequencing O-glycans from microgram quantities of glycoproteins. Anal Biochem. 2002;304(1):70-90.
- Royle L, Campbell MP, Radcliffe CM, et. al. HPLC-based analysis of serum N-glycans on a 96- well plate platform with dedicated database software. Anal Biochem. 2008;376(1):1-12.
- Walsh G. Biopharmaceutical benchmarks. Nature Biotechnology 2014;32(10):992–1000.
- Gong B, Burnina I, Stadheim TA, Li H. Glycosylation characterization of recombinant human erythropoietin produced in glycoengineered Pichia pastoris by mass spectrometry. J. Mass Spectrom. 2013;48(12):1308-17.
- Liu L, Gomathinayagam S, Hamuro L, et. al. The impact of glycosylation on the pharmacokinetics of a TNFR2:Fc fusion protein expressed in Glycoengineered Pichia Pastoris. Pharm Res. 2013;30(3):803-12.
- Tran BC, Barton C, Feng J, et. al. Glycosylation characterization of therapeutic mAbs by top- and middle-down mass spectrometry. Journal of Proteomics 2016;134(16):93-101.
- Zhang L, Luo S, Zhang B. Glycan analysis of therapeutic glycoproteins. MAbs. 2016;8(2):205- 15.
- Scott NE, Parker BL, Connolly AM, et. al. Simultaneous glycan-peptide characterization using hydrophilic interaction chromatography and parallel fragmentation by CID, higher energy collisional dissociation, and electron transfer dissociation MS applied to the N-linked glycoproteome of Campylobacter jejuni. Mol Cell Proteomics. 2011;10(2):201- 218.
- Hua S, Nwosu CC, Strum JS, at. al. Site-specific protein glycosylation analysis with glycan isomer differentiation. Anal Bioanal. Chem. 2012;403(5):1291-302.
- Hossler P, Khattak SF, Li ZJ. Optimal and consistent protein glycosylation in mammalian cell culture. Glycobiology 2009;19(9):936-49.
- Moh ES, Thaysen-Andersen M, Packer NH. Relative versus absolute quantitation in disease glycomics. Proteomics Clin. Appl. 2015;9(3-4):368-82.