The effects of different stress factors, i.e., heat, starvation, extreme pH, osmotic stress, antimicrobial agents, etc. on microorganisms recovered from manufacturing environments will be discussed. These factors common to manufacturing processes and drug product attributes have been argued as candidates for inclusion in study protocols to simulate stresses and will be discussed. The dynamic continuum from repairable cell damage, loss of cell viability to cellular death that microorganisms may experience contingent upon the pervading environment will be emphasized. The role of repair mechanisms addressing damage to cell membrane function, enzymatic activity, protein synthesis and nucleic acids will be reviewed in terms of the recovery and enumeration of microorganisms using growth and non-growth methods. We conclude that the use of stressed microorganisms as challenge organisms in method validation, suitability testing and growth promotion testing is impracticable and attempts to do so offer no scientific or quality value.
The current compendial methods are based on and qualified by the ability of viable microorganisms to grow on microbiological culture media forming visible colonies on solid media or exhibiting visible signs of microbial growth, e.g., turbidity, pellicle formation, precipitation, etc. in liquid media. Early in the development of microbiology as a scientific discipline, it was recognized that many viable microbial cells especially those in soil and water samples, do not proliferate in rich microbiological growth media (Staley and Konopka, 1985; Cundell, 2015). Recent work on the human microbiome has confirmed that our bodies contain numerous species not readily isolated by culture-based methods (Cundell, 2016). This limitation on the isolation of microorganisms due to their fastidious nature has not been fully appreciated by pharmaceutical microbiologists. This feature is genotypic in nature, i.e. governed by the limitations of the microorganisms biochemistry coded within their genome. Within inhospitable environments (such as arid cleanrooms absent of nutrients) microorganisms marshal their physiology using the ‘stringent response,’ a phenotypic characteristic. This includes the down-regulation of features required for growth and division, and up- regulation of stress response genes (Boutte and Crosson, 2016). The stress response essentially represents the trade-offs microorganisms make between growth, proliferation and a capacity to survive in these environments (Ferenci, 2016). In low nutrient environments cells can be regarded as existing in a ‘stationary’ phase with minimal or no replication, their physiology specifically geared toward survival (Morita, 1988; Hegge-Aronis, 2002; Hobbie and Hobbie, 2013). Cellular adaptations or stress responses assisting survival include thickening of the peptidoglycan layer, condensing of DNA, deactivation (but not destruction) of ribosomes, and reduction of cytoplasmic volume (Pietnev et al, 2015; Jaishanker and Srivastava, 2017).
USP General Informational Chapter <1223> Validation of Alternative Microbiological Methods that became official on December 1, 2015 contained a section discussing the limitations of the colony-forming unit (CFU) in terms of enumerating only those microorganisms that readily grow on solid microbiological culture media under the incubation conditions selected. The section highlights the inappropriateness of using the colony-forming unit as a gold standard for method validation when there are many surrogate markers (targeted signals) available other than the CFU for the detection of presence, and enumeration of microorganisms in water, air, pharmaceutical raw materials, drug substance and drug products.
The questions that this review article wishes to address are whether stressed microorganisms (i.e. those microorganisms exhibiting the well documented phenotypic phenomenon of stress responses) are practically feasible to generate, whether they are necessary challenges for growth-based microbial test methods, what stresses are appropriate for testing in support of pharmaceutical manufacturing and product release, and how stressed organisms might be artificially generated. With respect to alternative microbiological methods, the most suitable test standards will be proposed, given that the target signals are not colony-forming units. There is no consensus on the necessity to include in-house isolates, stressed micro- organisms and dead microorganisms as challenge organisms in method validation studies and whether the positions taken are merely recommendations or requirements (Table 1 and Table 2). Despite the absence of clear recommendations in FDA guidance documents for the use of in-house isolates, stressed microorganisms and dead microorganisms as challenge organisms in QC microbiology test- ing, several Form 483 observations have been cited during the inspection of pharmaceutical manufacturing facilities.

Stress Factors
In food microbiology, bacterial injury may be simply defined as the effect of one or more sublethal treatments on a microorganism. Often sublethal injury is the consequences of environmental insult such as chemical or physical processes that damages, but does not kill microorganisms (Bray, 1979; Wu, 2008; Wesche et al, 2009). Sublethal injury of microorganisms is distinct from microbial stress responses; the latter being a coordinated and directed expression of genes to phenotypically adapt to an inhospitable environment, such as reduced metabolism, dormancy, reduced cell size, and spore formation.
Within inhospitable environments, individual cells of populations do exhibit phenotypic heterogeneity in terms of stress responses. Such variance is advantageous to the survival of a population because it ensures that a subset of a microorganism will survive and later repopulate once conditions improve. This is a principle known as “bet-hedging” (Balaban et al, 2004; Kussell and Leibler, 2005; Geisel et al, 2011). Differences in stress tolerance among individual cells may therefore help by having a contingent of cells that can rapidly divide in times of plenty and by having a contingent of cells that can survive if conditions worsen.
Although microorganisms in a manufacturing facility exhibit stress responses in answer to environmental effects, it is almost entirely certain those unique phenotypes are swiftly lost in the laboratory. Upon experiencing an environment of a suitable nutrient composition facility isolates do not immediately proliferate, but transition through a period of latency (Monod, 1949). This latency period in which cell replication does not occur is commonly referred to as ‘lag phase’. During the lag phase, individual cells must alter their physiology from one geared toward assisting survival to one which supports replication. To begin dividing once more, microorganisms must alter their DNA superstructure, restructure cellular morphology, reorganize global metabolism and repair any oxidative damaged biomolecules encountered during stress response. Changes in a microorganism’s transcriptome and proteome between transitions from stationary phase (expressed during presence in the typical facility environment) to lag phase are significant with changes in the number and types of genes expressed. During transition from stationary to lag phase at least 75, 28, 39 genes in Bacillus licheniformis, Lactococcus lactis, and Salmonella enterica respectively are diff erentially expressed (Hornbaek et al, 2004; Larsen et al, 2006; Rolfe et al, 2009), Given the common themes reported, it is likely that any isolate recovered from the facility environment, transferred to an agar plate will rapidly and significantly alter their proteome and transcriptome, altering their features specifically adapted to survival in that environment to ones geared to replication and proliferation. Any features specifi c to a stressed physiological state are rapidly lost so there is no point of creating laboratory cultures of facility isolates.
Pin et al. (2009) have explained that old cultures employ a strategy of rejuvenation through replicative dilution. Cell division is asymmetrical in that one daughter cell retains old and damaged subcellular components (“old pole”), whereas the other daughter cell receives new components (“new pole”). As cells with new poles divide exponentially, old and damaged cells become serially diluted. In a similar fashion it is likely that damaged macromolecules and cellular structures are also diluted out to insignificance. Given that at least 105 viable cells are required for a visible colony of 0.2mm in diameter (Salvesen and Vadstein, 2000) a total of 18 cell divisions or generations are required. Clearly a colony of an isolate recovered from the facility is highly unlikely to possess environmental induced damage.
Upon transfer of microorganisms derived from a manufacturing environment in to laboratory conditions, modifications of important traits occurs, this has been termed “domestication” (Eydallin et al 2014). Steensels et al (2019) also reported on the phenomena of “domestication of industrial microbes”. They proved that during the domestication process, microbes gained the capacity to efficiently consume particular nutrients, cope with a multitude of industry- specific stress factors, often at the cost of a reduction in fitness in their original, natural environments, so, in time, during laboratory storage making them less representative of their original environments.
The above evidences that microorganisms encountering a stressful environment (e.g. low availability of water and nutrients) respond with the differential expression of genes generating a specific phenotype. Once cells are relieved of these conditions, cells (e.g. transferred to culture media) rapidly revert to the expression of a different set of genes. Therefore, the argument of applying facility isolates in to QC microbiology testing based upon the assumption they demonstrate stress response features is entirely without merit. Similarly, the argument of applying facility isolates in to QC microbiology testing after growth on culture media based upon the assumption they retain macromolecular and structural defects is entirely without merit.
The term stress has been used to describe the effect of sublethal treatments on the microbial cells (Bray, 1979; Wu, 2008; Wesche et al, 2009). Traditionally stress is measured by the difference in recovery on non-selective and selective media. A simple selective agent that can be added to a general microbiological growth media is salt, i.e., NaCl in the range of 0 to10%. Damage to the cell membrane usually results in an inability to make the transition to grow in selective media. This can be confirmed by propidium monoazide treatment, which only penetrates the microbial cells, if the cell membrane is damaged, complexing with DNA interfering with PCR amplification (Cangeiosi and Meschke, 2014). Representative physical and chemical sublethal treatment are presented in Table 3 a and 3 b.
Creation of Stressed Cells
Sublethal heat treatment and starvation are the simplest and most commonly used methods used to create stressed microbial cells. Table 4 contains the D Values and Z Values of common foodborne bacterial pathogens. In general, for vegetative cells Gram-negative bacteria are more heat susceptible than Gram-positive bacteria, and rods are more susceptible than cocci but there are exceptions.
The literature emphasize that moderate stress may result in a continuum of cellular injury that includes dead and surviving individuals. Within the survivors are injured and uninjured (normal) microbial cells (Kell et al, 1998; Davey, 2011; Cebrian et al, 2017). This means not all surviving cells will be injured (sublethal stress).
The effect of sublethal treatment on microbial cells include the following:
- Increased sensitivity to surface-active compounds, salts and toxic chemicals, antibiotics, dyes, and acid and basic pH.
- Loss of cellular materials
- Loss of cell membrane integrity
- Formation of endospores and smaller, dominant cells
- Longer lag phase when cultured
- Inability to multiply without cellular repair Location of the sublethal stress include the following:
Location of the sublethal stress include the following:
- Damage to cell surface structure, i.e., outer membrane in Gram- negative and teichoic acid in Gram-positive bacteria
- Damage to cytoplasmic membrane
- Damage to ribosomes
- Damage to DNA
- Damage to specific enzymes Repair (resuscitation) mechanisms
Repair (resuscitation) mechanisms
- Suspension in diluents, i.e., deionized water, buffers or peptone water
- Non-selective microbiological growth media (Soybean-casein digest broth)
- Media composition
- Specific agents in the media, e.g., pyruvate, EDTA, oxidase, etc.
- Incubation conditions, i.e., aerobic conditions, temperature, and duration
- Solid versus liquid media
- Stationary versus rotary incubation
- Time for repair
As microbial cells incubated in the presence of nutrients rapidly generate the cellular energy currency ATP using respiratory functions, mRNA that directs protein synthesis on ribosomes, and repairs cell membrane integrity, the resuscitation of microbial cells begins in minutes and usually is complete in a few hours (Ehrenberg, Bremer, Dennis, 2013). During a single generation time, which is around 20 minutes for E. coli grown in a rich medium, the bacterial cells have to duplicate a complete set of proteins, making up more than half of the cell dry weight and consuming around half of the available energy. Protein synthesis will be a trade-off between intracellular nutrition and energy costs.
Considerable attention has been given to the remarkable biosynthetic capabilities of E. coli in the biotechnology industry associated with the production of recombinant proteins. Neiβ et al (2019) calculated that approximately 13,500 ribosomes are required per cell at a growth rate of 0.7 per hour and 80% of ribosomes are actively translating, and the elongation rate is 16 amino acids per ribosome per second, then more than 170,000 amino acids per second are needed for protein synthesis.
The publication observed that this number equates to an individual tRNA turnover time of about 0.67 seconds. It is noteworthy that the cycle time comprises crucial steps such as the release of tRNA from ribosomes, transport for recharging at aminoacyl tRNA synthetases, and back transport to ribosomes. Millions of individual protein molecules are synthesized each generation. Although there is an evolutionary advantage for bacteria to reduce their generation time, as the rate of growth increases, the translation error rate will increase balancing this advantage.
Resuscitation steps employed in food, water and clinical microbiology differ between the necessary cellular repair that does not increase the cell number when used for enumeration and the enrichment that increases the cell number, to overcome an inherent limit of detection, for the detection of specific bacteria on selective media. For example, a resuscitation step prior to enumeration may be 1-3 hours in peptone water at 30-35°C while an enrichment step may be 18-24 hours in tryptic soy broth at 30-35°C. These are relative short incubation times compared to the total incubation time employed in compendial microbial test methods, e.g., at least 14 days for the <71> sterility tests.
Resuscitation Steps in Standard Microbial Test Methods
Resuscitation as a pre-incubation step is common in standard microbial test methods. In this section, examples will be given from food, water and pharmaceutical microbiology sectors, highlighting the short duration of the resuscitation steps.
As the foodborne bacterial pathogen Salmonella may be present in food in low numbers and in a debilitated (stressed) condition pre- enrichment in a non-selective medium overcomes these problems as well as that caused by adding large amounts of food to the selective enrichment broth. Tryptic Soy Broth incubated at 35°C for up to 24 hours is a common choice, followed by selective enrichment and isolation on diagnostic solid media (See Compendium of Methods for the Examination of Food). Modern microbiological methods may be employed in place of traditional methods. For example, a recent study demonstrated that a 24-hour pre-enrichment in buffered peptone water followed by a qPCR assay was found to be suitable for the detection of Salmonella spp. in low moisture foods (Wang et al, 2014).
For the enumeration of E. coli for chlorinated water, a neutralizing agent like sodium thiosulfate is added to the sampling bottle. The resuscitation of stressed or injured organisms is enhanced by initially culturing in an enriched, non-inhibitory media at a moderate temperature for a short period of time, e.g., tryptic soy broth at 35°C for 2-3 hours. The short pre-incubation period allows the resuscitation but not the proliferation of the indicator organism (See AWWA/APHA Microbiological Examination of Water 9212 B Recovery Enhancement).
Heterotrophic Plate Counts uses R2A agar for pour, spread, or membrane filter methods for monitoring the bacterial count in potable water. This low-nutrient agar gives higher counts than high- nutrient formulation. The highest counts are typically obtained with a 5 to 7 day incubation at a temperature of 20-25°C (See AWWA/APHA Microbiological Examination of Water 9215 Heterotrophic Plate Count).
USP <61> recommends the use of the general microbiological culture medium Soybean-Casein Digest Agar for microbial enumeration including the neutralizers Lecithin and Polysorbate 80 to overcome common antimicrobial preservative system in non- sterile drug products incubated at 30-35°C for 3-5 days for bacteria and 5-7 days for fungi. Method suitability testing is conducted with specified laboratory test strains at 30-35°C for NMT 3 days. The minimum incubation time of 3 days allows for the resuscitation and enumeration of stressed bacteria.
Experience with Alternate Methods to USP <71> Sterility Tests
USP <71> Sterility Tests requires the use of Fluid Thioglycollate and Soybean-Casein Digest Media incubated aerobically at 30-35°C and 20-25°C respectively for at least 14 day for sterility testing. The incubation time was extended from 7 to 14 days based on the report by Bathgate et al (1993) that showed that in greater than 30% of the failing tests the organisms, presumably stressed, exhibited visible growth between the 7th and 14th day of incubation. Although microorganisms detected in <71> are likely to be stressed, the question can be asked why was so long an incubation time required to detect microbial growth in the sterility test broth? Possible reasons include suboptimal media (Fluid Thioglycollate Medium), suboptimal incubation temperatures (20-25°C), high number of microbial cells needed to result in obvious turbidity (the order of 108 CFU/mL), and injured microbial cells in the test specimens. Two other factors that bear consideration for the delayed growth in sterility test broth are reduced quorum sensing with very low densities of microbial cells in liquid culture and the formation of dominant cells that are a survival strategy in the face of adverse conditions.
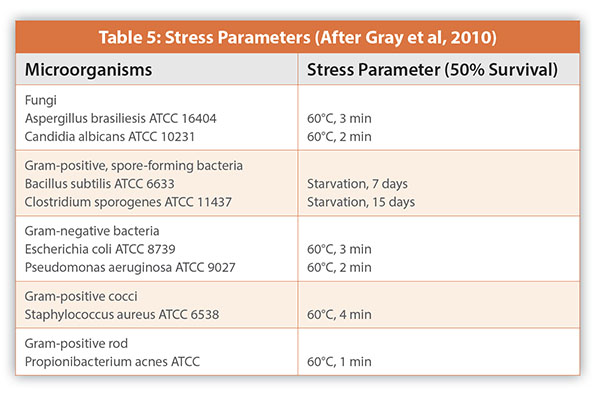
As a preliminary study to the development and validation of the Milliflex™ System as an alternative method to the <71> sterility test, Gray et al (2010) evaluated ten different solid agar media and three incubation conditions using twenty-two difference unstressed and stressed Quality Control organisms and in-house isolates (Table 5). The authors questions the use of sublethal heat treatment and starvation for exposure time to achieve a 50% reduction on cell count as a method in generating stressed microbial cells. The treated cells are likely to be a continuum of live, damaged, and dead cells not just stressed cells. This may be highlighted in Figure 4 showing heat treated and untreated Kocuria rhizophilia colonies on tryptic soy agar incubated at 30-35°C for three days. The untreated colonies are of uniform size and shape while the treated colonies varied in size and shape. As expected, the sublethal stress increased the lag phase by one to three hours but this will not be significant with incubation times numbered in days and not hours. Schaedler blood agar incubated at three difference conditions (aerobically at 20-25°C, aerobically at 30-35°C and anaerobically at 30-35°C) gave the greatest recovery of strict aerobes, facultative anaerobes, and strict anaerobes.
Not only is the use of stress and dead cells in method validation unnecessary, in the absence of a consensus of what is a standardized protocol, their use cannot be consistently applied. Within the pharmaceutical industry, furthermore, the starvation of spore-forming microorganisms in distilled water for up to 15 days may reduce the proportion of vegetative cells in the inocula, but will not stress resistant spores.
Rapid sterility tests may be subdivided into growth-based and non- growth-based methods. These methods differ by target signal from microbial growth, enzymatic activity to vital staining to nucleic acid amplification (Table 6). Standard materials suitable for these tests methods may range from viable microorganisms, ATP, dead fluores- cent bacterial cells, and genomic materials.
Conclusions
The resuscitation time required for a sublethally stressed microbial cell to enter the log phase of growth is measured in hours while the incubation time for microbial test methods is measured in days, making it unnecessary to use stressed cells for method validation and growth promotion and method suitability testing. Standard industry protocols are lacking for the creation of both stressed and dead cells making their use unreliable. Furthermore, as recommended in USP <1223> Validation of Alternate Microbiological Tests - Performance Acceptance option it is not necessary to demonstrate that an alternative method is comparable to the compendial method, especially when the targeted signal is not a viable microorganism measured as a colony-forming unit.
References
- Balaban N. Q., Merrin J, Chait R, Kowalik L, Leibler S. 2004. Bacterial persistence as a phenotypic switch. Science 305:1622–1625. https://doi.org/10.1126/science.1099390
- Boutte, C.C., and S. Crosson 2016 Bacterial lifestyle shapes stringent response activation. Trend in Microbiol. 21 (4):174-180.
- Bray, B. 1979 Methods to detect stressed microorganisms. J. Food Prot. 42(4): 340-355
- Cangeiosi, G. A., and J.S. Meschke, 2014. Dead or alive: Molecular assessment of microbial viability. Appl. Environ. Microbiol. 80(19): 5884-5891
- Cebrian, G., S. Condon, and P. Manas 2017 Physiology of the inactivation of vegetative bacteria by thermal treatment: Mode of action, influence of environmental factor and inactivation kinetics Foods 6(107): 1-21
- Cundell, T. 2015 The Limitations of the Colony-forming Unit in Microbiology. Eur. Pharm. Rev. 20(6):11-14
- Cundell, A.M. Microb. Ecol. Microbial ecology of the Human Skin https://doi.org/10.1007/ s00248-016-0789-6 2016
- Davey, H. M. 2011 Life, death, and in-between: Meanings and methods in microbiology. Appl. Environ. Microbiol. 77:5571-5576
- Eydalllin, G., B. Ryall, R. Maharjan and T. Ferenci. (2014), “The nature of laboratory domestication changes in freshly isolated Escherichia coli strains”, Environmental Microbiology 16(3), 813-828
- Gray, J. C., A. Staerk et al, 2010 Growth-promoting properties of different nutrient media evaluated with stressed and unstressed microorganisms: Pre-study for the validation of a rapid sterility test. PDA J. Pharm. Sci. & Technol. 64(3): 249-263
- Geisel N, Vilar JMG, Rubi JM. 2011. Optimal resting-growth strategies of microbial populations in fluctuating environments. PLoS One 6:e18622. https://doi.org/10.1371/ journal.pone.0018622.
- Hobbie, J. E, and E.A. Hobbie 2013. Microbes in nature are limited by carbon and energy: the starving-survival lifestyle in soil and consequences for estimating microbial rates. Front Microbiol 4:324. https://doi.org/10.3389/fmicb.2013.00324
- Hornbaek T, Jakobsen M, Dynesen J, Nielsen AK. 2004. Global transcription profiles and intracellular pH regulation measured in Bacillus licheniformis upon external pH upshifts. Arch Microbiol 182:467– 474. https:// doi.org/10.1007/s00203-004-0729-6.
- Jaishankar J, Srivastava P. 2017. Molecular basis of stationary phase survival and applications. Front Microbiol 8:2000. https://doi.org/10 .3389/fmicb.2017.02000.
- Kell, D.B., A.S. Kaprelyants, et al 1998 Viability and activity in readily culturable bacteria: a review and discussion of the practical issues. Antonio van Leeuwenhoek 73: 169-187
- Kussell E, Leibler S. 2005. Phenotypic diversity, population growth, and information on fluctuating environments. Science 309:2075–2078. https://doi.org/10.1126/ science.1114383
- Monod J. 1949. The growth of bacterial cultures. Annu Rev Microbiol 3:371–394. https:// doi.org/10.1146/annurev.mi.03.100149.002103
- Larsen N, Boye M, Siegumfeldt H, Jakobsen M. 2006. Differential expression of proteins and genes in the lag phase of Lactococcus lactis subsp. lactis grown in synthetic medium and reconstituted skim milk. Appl Environ Microbiol 72:1173–1179. https://doi.org/10.1128/ AEM.72.2.1173-1179.2006.
- Morita, R. Y. 1988. Bioavailability of energy and its relationship to growth and starvation survival in nature. Can J Microbiol 34:436–441.
- Nieβ , A., M. Siemann-Herzberg and R. Takors 2019 Protein production in Esherichia coli is guided by the trade-off between intracellular substrate availability and energy cost. BMC Microb. Cell Fact. 18:8 1-10
- Pin C, Rolfe MD, Munoz-Cuevas M, Hinton JCD, Peck MW, Walton NJ, Baranyi J. 2009. Network analysis of the transcriptional pattern of young and old cells of Escherichia coli during lag phase. BMC Syst Biol 3:108. https://doi.org/10.1186/1752-0509-3-108.
- Pletnev P, Osterman I, Sergiev P, Bogdanov A, Dontsova O. 2015. Survival guide: Escherichia coli in the stationary phase. Acta Naturae 7:22–33
- Rolfe MD, Rice CJ, Lucchini S, Pin C, Thompson A, Cameron ADS, Alston M, Stringer MF, Betts RP, Baranyi J, Peck MW, Hinton JCD. 2012. Lag phase is a distinct growth phase that prepares bacteria for exponential growth and involves transient metal accumulation. J Bacteriol 194: 686–701. https://doi.org/10.1128/JB.06112-11
- Salvesen, I.; Vadstein, O 2000. Evaluation of plate count methods for the determination of maximum specific growth rate in mixed microbial communities, and its possible application for diversity assessment. J. Appl. Microbiol. 88 (3), 442–448
- Staley J.T., Konopka A. Measurement of in situ activities of nonphotosynthetic microorganisms in aquatic and terrestrial habitats. Annu Rev Microbiol. 1985;39:321-46. doi: 10.1146/annurev.mi.39.100185.001541. PMID: 3904603
- Steensels, J., B. Gallone, K. Voordeckers, and K. Verstrepen. (2019), “Domestication of Industrial Microbes,” Current Biology 29, R381-R-393
- Wang, H., V.S. Gill, et al 2014 Evaluation and comparison of rapid methods for the detection of Salmonella in naturally contaminated pine nuts using different pre enrichment media. Food Microbiol. 46: 58-65
- Wesche, A. , J. B. Gurtler et al, 2009 Stress, sublethal injury, resuscitation, and virulence of bacterial foodborne pathogens. J. Food Prot. 72(5): 1121-1138
- Wu, V. C. H. 2008 A review of microbial injury and recovery methods in food. Food Microbiol. 25: 735-744
- Måns Ehrenberg, Hans Bremer, Patrick P. Dennis, Medium-dependent control of the bacterial growth rate, Biochimie, Volume 95, Issue 4, 2013, Pages 643-658, ISSN 0300-9084