Introduction
Since the inception of Limulus amebocyte lysate (LAL) testing [1], the past four decades of testing have proven fruitful in terms of making in vitro predictions of in vivo responses regarding the significant pyrogen of drug manufacturing, bacterial endotoxin. The Limulus test has served as the basis of preclusion of bacterial endotoxin in drug products for these many years. However, current discoveries elucidating the mechanisms by which endotoxin interacts with host biosensors1 in both invertebrates and vertebrates is turning many a preconceived notion on its head. Today’s researchers have found that both endotoxins and their biosensors are much more dynamic structures than previously believed. The dynamism for both arises from the genomes of bacterial and metazoan host respectively. For example,
- some lipopolysaccharide (LPS) variants are silent or antagonistic (opposite agonistic expectations)
- some host sensor responses do not agree among closelyrelated species (sometimes caused by single amino acid substitutions in the biosensor or accessory proteins)
- individual polymorphisms within a given species (including humans) render some individuals oblivious to endotoxin
- counter-intuitively, some pathogens are pathogenic because they do not elicit a host response via Toll-like receptor 4 (TLR4)
- whereas many mammalian symbionts elicit strong deleterious host responses via TLR4
- host compartmentalization of biosensors in different mammalian tissue types produces a gradient of available TLR4 and accessory molecules that define custom host responses
To begin to make some broad sense of various crosscurrents, we need to examine the evolutionary basis of endotoxin biosensors via an ancient-to-modern perspective. Given the mass of data accumulating, and the number of potential tests on the horizon (Elisa [2], whole blood [3], non-LAL protein binding [4], lab on a chip [5], novel mechanistic [6, 7] physical parameter methods [8-10] etc.), it is good to examine our prejudices regarding what we expect an endotoxin biosensor to tell us from a parenteral preclusion vantage. Is it best to detect (a) as many pieces and parts of Gram-negative artifacts as possible (assorted lipoprotein, LPS and LPS-like molecules), (b) to detect only those artifacts perceived to be specifically or particularly deleterious to human hosts or (c) is it prudent to tread a median path that has some elements of both approaches?
Endotoxin Biosensor Approaches Used in Nature
A survey of ancient invertebrate and modern mammalian biosensor approaches used to detect microbial invaders (contaminants) reveals a number of strategies, some overlapping. The first may simply be called a generic approach that identifies friend and foe based upon the general pattern of LPS core and lipid A acyl chains attached; it is an ancient, and to some degree, conserved approach used in modern systems with increasing specificity. The most basic approach uses defined sequences of Leucine rich repeats (LRR) which occur in amphiphilic loops to literally snag hydrophobic sequences of pathogen-associated molecular patterns (PAMPS) such as LPS. The best-studied example of this is Limulus or Carcinoscorpius Factor C molecule, the active biosensor that serves as the basis of the LAL test reaction. The modern approach, as shared by mammalian hosts, is an accumulation of predilections that makes use of the ancient LRR pattern recognition receptor (PRR). The modern approach uses some elements of the ancient machinery but has separated it into a number of complex biosensors woven around an ancient LRR base. Additionally, we will see that in modern blood and tissue there are recognition distinctions based upon where infection occurs within the body (some locations having numerous receptors and some having few or no receptors). This modern attempt at localization seeks to avoid the systemic spread of infection just as Limulus attempts to wall offinvaders in primitive fashion with its Factor C-initiated blood coagulation defense.
Endotoxin or LPS, and more specifically the lipid A end, is a potent PAMP and exists as a structural component of the Gramnegative bacterial outer membrane. Its unique architecture serves to distinguish its chemical structure from all other microbial contaminants. However, the degree of recognition by the host varies with the LPS type (as genetically encoded by various gram negative bacteria (GNB)) and/or lipid A types produced. The prototypical PAMP is E. coli biphosphorylated and hexa-acylated LPS. Endotoxins or Lipid A analogs that are variants of the prototypical structure (monophosphorylated on either end of the glucosamine backbone, containing fewer or greater number of acyl chains or variant numbers of carbons per acyl chain, as well as variations in the KDO core sugar (3-deoxy-D-manno-octulosonic acid) will typically diminish the host response. Many studies have been done historically deciphering the nuances of responses by variant LPS and lipid A structures relative to the resulting activation of rabbit pyrogen, LAL, and cytokine production, such as in whole blood assays. [11]
Some hosts do not mount an alarm or defense in the presence of LPS containing certain fine structural features. In this way, each modern species of mammal has an “endotoxic-principle” that has been evolutionarily developed and which varies slightly as to the type of bacterial endotoxin that it recognizes. For example, lipid IVa is a precursor to lipid A in the process of bacterial biosynthesis and, when presented in two different species of mammals, mice and humans, each mammal elicits a different host response. Mice recognize and mobilize a response to lipid IVa whereas humans do not (the “antagonistic activity of lipid IVa is determined by human MD-2” [12]). The species specific nature of mammalian response contrasts with primitive metazoan responses (such as Limulus’) which utilize a more generic (presence-absence) approach to fighting GNB invaders. For example, Limulus responds to tetra, tri, and penta-acyl lipid A as vigorously as it does to hexa-acyl lipid A which is not generally the case in mammals. [11]
An Evolutionary-based Perspective
A comparison of ancient Factor C to modern biosensor function as represented by the TLR4/MD2 dimer complex is instructive since both Limulus and modern mammalian innate immune systems appear primarily concerned with systemic dissemination of E. coli type LPS, as sensed by both Factor C and TLR4/MD2, respectively. In both systems this makes a good deal of sense as Limulus has no way outside of coagulation to partition off its blood system and mammals must be concerned as they harbor symbiotically large reservoirs of helpful bacteria that could turn deadly very quickly. After all, the only difference between a live host and a dead host in regard to containing populations of GNB is the ability to keep them confined to the desired reservoirs and feeding upon desired metabolites.
Since any metazoan cannot survive long without mechanisms to detect endotoxin [13] and neutralize Gram-negative invaders, then these mechanisms are highly conserved, and we cannot conceive that between mammals and arthropods that there could be significant lapses in organism types that possess such mechanisms. While such a system must be conserved to promote survival, we can expect that workarounds and parallel systems would develop via mutation and that, eventually, the latter might supplant the former. The mechanisms that now exist in vertebrates and mammals are highly diverged from the ancient mechanisms found to still reside in modern day invertebrates including Limulus as a “living fossil”. However, even though the detection means have diverged, the end goal of the detection of endotoxin, the specific forms responded to, the wide spectrum of that response, and the associated organism types detected remains similar in ancient and modern systems. The commonly accepted premise underlying these facts can be stated simply: Current methods used by vertebrates are derived from a multitude of simpler systems inherited via a common ancestor of vertebrates and invertebrates [14]. Additionally, homology in Lipid A-sensing-remnants (plural) maintains the similarity of host responses to endotoxin detection via biosensors despite the wide divergence of host physiology.
Such conservation of necessary biosensor gene sequences should not come as a shock as many of the most basal biological systems including ribosomes [15] and Hox genes2 [16] have been preserved from the most distant primordial ancestors. Also, some very primitive organisms possess complex genetic “toolkits” [17]. While modern systems of innate immunity have grown in complexity via mutation and duplication (with added functionality), they have also retained their most basic function: the ability to sense via PRRs the unique geometry of Lipid A as a PAMP. Figure 1 shows how one primitive system for the detection of endotoxin can be both preserved and supplanted over time by the development of parallel systems. Once analogous biosensors are in place, the prototypical system is free to degrade partly or wholly (partly in that it can complement the new system by retaining some reduced functionality or cofunctionality).
 Figure 1. Hypothetical evolution of endotoxin biosensors, ancient to modern. Workarounds evolved from an original, simple system, the prototypical system (ancient at left), into a variety of parallel, interacting responses that are more complex as built around the original system. Loops shown symbolize Leucine Rich Repeats (LRR) typical to endotoxin biosensors. Biosensor at left can be viewed as analogous to Factor C (Limulus) and the more complex interplay of biosensor and accessories at right can be viewed as analogous to TLR4 /MD-2/ CD-14/LBP (lipopolysaccharide binding protein).
Figure 1. Hypothetical evolution of endotoxin biosensors, ancient to modern. Workarounds evolved from an original, simple system, the prototypical system (ancient at left), into a variety of parallel, interacting responses that are more complex as built around the original system. Loops shown symbolize Leucine Rich Repeats (LRR) typical to endotoxin biosensors. Biosensor at left can be viewed as analogous to Factor C (Limulus) and the more complex interplay of biosensor and accessories at right can be viewed as analogous to TLR4 /MD-2/ CD-14/LBP (lipopolysaccharide binding protein).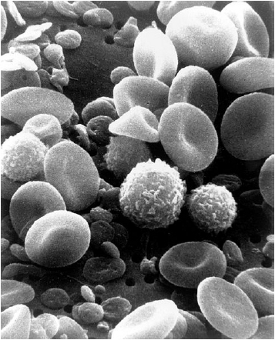 Figure 2. Wide diversity of modern blood cells contrasts Limulus’ single blood cell (amebocyte).Public domain (U.S. Government).
Figure 2. Wide diversity of modern blood cells contrasts Limulus’ single blood cell (amebocyte).Public domain (U.S. Government).A similar process of differentiation can be seen in the growing complexity of blood systems, ancient to modern, in that the single circulating cell of Limulus (amebocyte) has become a large number of specialized cells (Figure 2) producing a huge number of proteins possessing various and sophisticated functions in vertebrates. The progression of biosensor differentiation can be seen to have occurred in nature when we view some representative systems side by side, including those of Limulus, Drosophila (ancient), and mammals (modern) in Figure 3. The Drosophila pattern (Figure 3, center) demonstrates how modern TLRs may have been co-opted as the toll in the fly serves in patterning the body plan. Both ancient and modern systems rely upon recognizing the geometry (which can be viewed as a sum of molecular charges and hydrophobic-hydrophilic effects with a resulting 3-dimensional structure) of the lipid A end of the LPS molecule.3 The most potent host responses occur presumably because the types of organisms harboring the prototypical structures belong to bacteria perceived as most dangerous from a systemic vantage and therefore the biosensor-associated response provides the most survival utility to its possessor. Meanwhile, on the inside of the cell (post-PAMP recognition via PRRs) the resultant response to transmembrane signaling events (ancient or modern) proceed by ancient, conserved mechanisms of intracellular signaling including the NF-kb pathway [18] ending in a message to the cell nucleus to manufacture various defensins (ancient) and/or cytokines (modern) via the expression of specifi c genes.
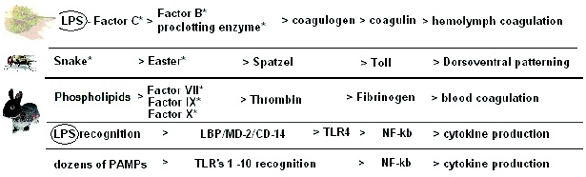 Figure 3. Proteolytic cascades of Drosophila, Limulus, and Mammals occurring in the hemolymph/ blood and resulting in coagulation and Toll type pathway activation. The two at the bottom are simplifi ed views of TLR4 and TLR1-TLR10 signaling respectively. Adapted from several references including: A Toll-like receptor in horseshoe crabs, Immunological Reviews 198/2004 (pg. 107). Note that Limulus ancestors are thought to predate all insects by at least 50 million years (<400 [19] versus ~450 MY [20]). *denotes serine protease zyomgens [21].
Figure 3. Proteolytic cascades of Drosophila, Limulus, and Mammals occurring in the hemolymph/ blood and resulting in coagulation and Toll type pathway activation. The two at the bottom are simplifi ed views of TLR4 and TLR1-TLR10 signaling respectively. Adapted from several references including: A Toll-like receptor in horseshoe crabs, Immunological Reviews 198/2004 (pg. 107). Note that Limulus ancestors are thought to predate all insects by at least 50 million years (<400 [19] versus ~450 MY [20]). *denotes serine protease zyomgens [21].Factor C – an ancient biosensor
Multifunction
In order to compare ancient and modern systems, we need some knowledge of Factor C’s (Limulus, Carcinoscorpius, Tachypleus) multifunctionality and structure. Serine protease cascades are widely used in innate immunity and occur in many variant forms. The broad functionality of Factor C as the biosensor of Xiphosura can be seen by viewing its many combined functions in contrast to the use of differentiated molecules in modern systems. The multi-functionality of Factor C can be summarized as serving the following diverse functions in the invertebrate: biosensor, coagulation zymogen with complement like activity [22], opsonization of bacteria [23, 24], and as a hemocyte receptor [25].
The functional characterization of Factor C and its eventual commercialization can be studied by looking at three patents, two by Jeak L. Ding, Bow Ho and Nguan S. Tan of the National University of Singapore: US 5,716,834 (Feb 10, 1998) and US 6,719,973 B1 (Apr 13, 2004), and a third by Jeak L. Ding and Bow Ho, US 7,763,704 B2 (Jul 27, 2010). These patents serve to illuminate the utility of this single invertebrate molecule as: (a) providing the biosensor Factor C (rFC), (b) providing cDNA probes that allow the search for similar sequences in different source organisms or tissue types, (c) providing strong LPS binding sequences to remove endotoxin from solutions in drug manufacturing or research processes, (d) providing potential anti-LPS (Sepsis) therapeutic drug candidates, and (e) helping to determine the defi nitive endotoxin binding sequences in ancient and modern systems. Tan, Ho, and Ding discuss the difficulty of cloning and producing Factor C. “The difficulty in doing so was largely due to its highly complex mosaic structure. While many highly disulfi debonded proteins have been successfully expressed...few display the kind of complexity posed by the Factor C LPS binding domain” [26].
An interesting observation by Tanaka [27] includes the apparent rationale for the necessity of different biosensor functionality in widely divergent body types (ancient invertebrates versus more modern vertebrates):
One may ask that if the final step of hemostasis has been so simple and effective for hundreds of million years, what is the advantage of having multiple coagulation Factors and complex pathways to achieve the same goal (clotting) as in invertebrates? Among nonmammalian and mammalian vertebrates, amino-acid sequences of prothrombin and fibrinogen are well conserved. The important evolutionary difference between the vertebral coagulation system and that of invertebrate species is the need to provide localized thrombosis in high pressured closed networks of blood vessels in contrast to the low-pressure open circulation.
Structure
The use of Leucine-rich repeats (LRR) by metazoans to snag bacteria and other microorganisms and their artifacts4 is very old, dating back probably 2 billion years and existing today also in plants [28]. There are many such natural molecules that have been found that bind microbial PAMPs, including some that specifically bind endotoxin. The general pattern necessary to bind endotoxin effectively is described by Frecer, Ho and Ding [29], and shown to consist of a minimum amphipathic structural motif of basic (B), polar (P) and hydrophobic (H) amino acid residues in either β-sheet or β-hairpin loop conformation.
… the minimum amphipathic lipid A- binding sequence (is) BHPHB of fi ve alternating basic, polar and hydrophobic residues with β-sheet backbone conformation... However, less symmetrical patterns containing various arrangements of two or more basic residues within a motif of 5±7 amino acids were found in all the considered sequences (they studied). Patterns with a higher content of basic residues BHBHB showed a lower number of hits.
Awareness of the general LRR rule helps illustrate what some have proposed as a more specifi c rule when it comes to Factor C binding of endotoxin. Factor C has strong LPS binding sites necessary to complement its biosensor function at the serine protease end of the molecule, and since LPS Lipid A analogs serve to defi ne the essence of both TLR4 and Factor C biosensor response to various degrees, then we know that both TLR4 and Factor C interact with LPS directly (with and without accessory helpers, respectively). A “tripeptide motif” model has been proposed by Koshiba et al [30] who showed that without the specifi c tripeptide sequences in the Cys-rich region of CrFC, there would be no binding and subsequent reaction with endotoxin.
We propose that the fi rst conserved tripeptide motif (Arg36- Trp37-Arg38) in the Cys-rich region of Factor C plays an important role in the recognition of LPS, an assumption that is consistent with a modeling study of ALF5 that predicts direct docking of the corresponding tripeptide motif with LPS. In this model, two lysine residues that correspond to Arg36 and Arg38 of Factor C interact with glucosamine (GlcN) I-1-phosphate in lipid A, and a tryptophan residue, which corresponds to Trp 37 of Factor C, interacts with a hydrophobic portion of lipid A. Our results are consistent with such a model and underscore the importance of these elements within the Cys-rich region of Factor C for mediating its association with LPS, an essential structural characteristic that appears to be conserved among other LPS-recognizing proteins.
Koshiba et al. displayed sequences for four modern proteins (three human and a rabbit) next to two ancient proteins (Factor C and anti- LPS factor from Limulus) which give us good examples of the lineup of critical, ancient to modern, amino acids in sequences shared in such LPS recognition and binding-type proteins (Figure 5). This concept has also recently been used to devise analogs that might be competitive at blocking the biosensor-LPS interaction, thus ameliorating the deleterious host response in various inflammations [31] and in sepsis. Furthermore, these sequence binding determinants might give us a way to see where in modern systems the ancient biosensor sequences now reside.
 Figure 5. Conserved tripeptide motifs in Factor C are critical for LPS interaction. The sequence of Factor C includes binding regions that precede the Sushi regions shown previously. Two of the tripeptide motifs in the Cys-rich region (36–38 and 55–57) are underlined as well as the LPS-recognizing sites of Sushi peptides 1 and 3 and fi ve additional proteins. The amino acid sequences are numbered according to positions in the mature proteins. hLBP, humanLPS-binding protein; hBPI,human bactericidal permeability-increasing protein; hMD-2, human MD-2; lALF, Limulus anti-LPS factor; and rCAP18, rabbit cationic antimicrobial protein. Derived from Koshiba et al [29].
Figure 5. Conserved tripeptide motifs in Factor C are critical for LPS interaction. The sequence of Factor C includes binding regions that precede the Sushi regions shown previously. Two of the tripeptide motifs in the Cys-rich region (36–38 and 55–57) are underlined as well as the LPS-recognizing sites of Sushi peptides 1 and 3 and fi ve additional proteins. The amino acid sequences are numbered according to positions in the mature proteins. hLBP, humanLPS-binding protein; hBPI,human bactericidal permeability-increasing protein; hMD-2, human MD-2; lALF, Limulus anti-LPS factor; and rCAP18, rabbit cationic antimicrobial protein. Derived from Koshiba et al [29].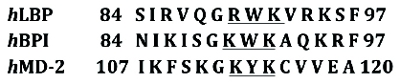 Figure 6. Human LBP, BPI and MD-2 share the key tri-peptide binding motifs prototypical of the ancient endotoxin biosensor (Factor C). Importantly, it has been shown that the relevant amino acids forming the “tripeptide motifs” line up with endotoxin binding regions in the complex three dimensional structures (it would be of little relevance if the linear sequence matched the motif but did not line up in the 3-D structure).
Figure 6. Human LBP, BPI and MD-2 share the key tri-peptide binding motifs prototypical of the ancient endotoxin biosensor (Factor C). Importantly, it has been shown that the relevant amino acids forming the “tripeptide motifs” line up with endotoxin binding regions in the complex three dimensional structures (it would be of little relevance if the linear sequence matched the motif but did not line up in the 3-D structure).Given the discovery of these binding site motifs, it is becoming apparent that there remain unrealized areas of endotoxin interaction in the blood system that includes binding by blood proteins such as hemoglobin [32] that are not currently considered to be a signifi cant part of the host response to endotoxin [33]. Differentiating the body’s response and the effects of LPS via non-receptor means may be under-defined but the involvement of the complement system via mechanisms that do not include TLR4 seem certain [34].
Modern endotoxin biosensor mechanisms
Since the tripeptide motifs shown in Figure 4 are shared among both ancient and modern endotoxin biosensors, there is reason to believe this mechanism has been widely passed down ancient to modern (rather than developed independently). Several of the peptides shown in Figure 5 have more than one overlapping tripeptide motif. This tripeptide motif – a subset of LRRs in general – exists in several places in the Factor C molecule and seems to be split into many places in the mechanism of the modern mammalian biosensor as existing in TLR4, MD-2, LBP and BPI. This modern interaction via a complex group of parts, (contrasting with the ancient Factor C biosensor in which many motifs occupy the same molecule), culminates in pattern recognition via a receptor called the “Toll-like Receptor” (TLR) on the one hand and also the activation of a set of blood coagulation factors called “complement” on the other.
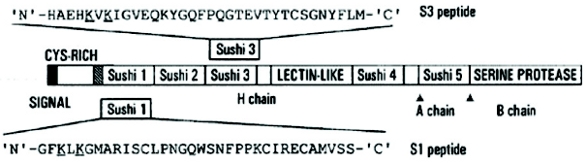 Figure 4. Factor C [rFC is 994 amino acids (heavy and light chains)] and its derived Sushi 1 and Sushi 3 peptides. The truncated fragments of the CrFC* protein (CrFCES), S1 and S3 peptide amino acids are listed and the endotoxin binding regions are underlined. Adapted from patent 6,719,973 B1 (Apr 13, 2004). *Cr is Carcinoscorpius and FC is Factor C.
Figure 4. Factor C [rFC is 994 amino acids (heavy and light chains)] and its derived Sushi 1 and Sushi 3 peptides. The truncated fragments of the CrFC* protein (CrFCES), S1 and S3 peptide amino acids are listed and the endotoxin binding regions are underlined. Adapted from patent 6,719,973 B1 (Apr 13, 2004). *Cr is Carcinoscorpius and FC is Factor C.Toll was first identifi ed in Drosophila6 [35] during studies on axial patterning and subsequently similar structures were found in higher animals and termed “Toll-like”. Both arms of higher animal innate immunity have analogs in Factor C. It is as if the ancient Factor C were splintered eons ago (last common ancestor which was arguably some kind of worm) and now resides in many different proteins spanning two different arms of innate immunity (complement and TLR4 activation). Including all rFC binding region sequences above expands the tripeptide motif to RWK, KWK, KYK, KLK, KFR and KVK. Using a genetic code table we can see that lysine, K, and arginine, R, are easily substituted in some instances by a single amino acid change: Arg=R is formed by codons AGA and AGG which can be mutated to Lys=K, AAA and AAG, and thus the R or K/W or Y/R or K “tripeptide motif” appears to loosely defi ne what may have come down via the endotoxin biosensor in terms of the ancient mechanism. From a combination of the structure of Factor C cys-rich region and that of Sushi 1 and Sushi 3, we get the following possible sequences for Factor-C-like binding sites in LPS biosensors: [R / K] [W, Y, L, F or V] [R / K]. The number of binding motifs and their 3-dimensional arrangement (β-sheets and β-hairpin loops being prototypical) in the folded molecule are the determinants of endotoxin binding.7 This knowledge better equips us to make a judgment as to whether the current vertebrate Toll-like machinery (TLR-4 and accessories LBP, MD-2 and CD-14) are scattered remnants of this ancient system.
Many key pieces of the modern biosensor machinery CD-14, TLR4, LBP, and MD-2, contain LRRs, the latter three are known to contain specifi c endotoxin sequences and CD-14 has become necessary to complete the biosensor function of MD-2/TRL-4. Since CD-14 is present in both soluble form (blood) and membrane bound form (without a transmembrane signaling function in man and mice) it reminds one of Factor C in this regard (soluble (s) and hemocyte membrane (m) bound forms). An excerpt from Palsson-McDermott et al [36] summarizes the early phase recognition of endotoxin by LBP/CD-14:
Probably the first host protein involved in LPS recognition is LPSbinding protein (LBP). LBP is an acute-phase protein, produced in the liver, which circulates in the bloodstream where it recognizes and forms a high-affinity complex with the lipid A moiety of LPS, as free molecules, fragments, or still bound to the outer membrane of intact bacteria... The role of LBP appears to be that of aiding LPS to dock at the LPS receptor complex by initially binding LPS and then forming a ternary complex with CD14, thus enabling LPS to be transferred to the LPS receptor complex composed of Toll-like receptor-4 (TLR4) and MD-2. CD14 is found in two forms. The fi rst is soluble CD14 (sCD14), which occurs in plasma where it helps to convey LPS signaling in cells lacking membrane-bound CD14, e.g. endothelial and epithelial cells. The second, more extensively studied form of CD14, is membrane bound (mCD14), attached to the surface of myeloid cells via a glycosylphosphatidyinositol tail, enabling CD14 to be membrane proximal despite lacking a transmembrane domain.
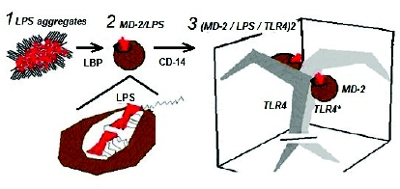 Figure 7. Cartoon of modern host biosensor machinery. (1) LBP extracts LPS monomer from aggregate and facilitates transfer into (2) MD-2 structure with all but one of the hexa-acyl chains fi tting inside the MD-2 hydrophobic-lined pocket or “basket” and the sixth acyl chain (E. coli) protruding to support binding to TLR4* - the second pair of the TLR4 dimer. (3) A side view of the TLR4 dimer (MD-2/LPS/TLR4)2. Transmembrane structure is shown below the plane of the cell membrane.
Figure 7. Cartoon of modern host biosensor machinery. (1) LBP extracts LPS monomer from aggregate and facilitates transfer into (2) MD-2 structure with all but one of the hexa-acyl chains fi tting inside the MD-2 hydrophobic-lined pocket or “basket” and the sixth acyl chain (E. coli) protruding to support binding to TLR4* - the second pair of the TLR4 dimer. (3) A side view of the TLR4 dimer (MD-2/LPS/TLR4)2. Transmembrane structure is shown below the plane of the cell membrane.Adding to the complexity of host biosensor detection of endotoxin, CD-14 has been found to allow TLR4 binding of smooth LPS (O-chain containing isoforms), whereas rough LPS does not require CD-14. The knowledge of this distinction was brought about by the Heedless mutation that showed a premature stop codon inserted into the CD- 14 gene (in mice) [37]. The existence of such a mutation (stop codon insertion) points to the hypothetical model shown in Figure 1, in which biosensors are shown to be in flux, evolutionarily speaking, and change in functionality over time. The key piece of the modern mechanism that engulfs LPS is MD-2. Nobel Prize winner (2011) Bruce Beutler called MD-2 a “basket-like” structure; indeed, it takes upon itself a woven appearance in molecular models and is a “basket” that comes to contain Lipid A in an internal hydrophobic pocket. It then deposits itself with LPS into the dual spaces that form the TLR-4 dimer. The cartoon in Figure 7 is loosely-based upon figures by Park et al [38].
Fine-tuning endotoxin recognition To partition intruders via modern biosensors
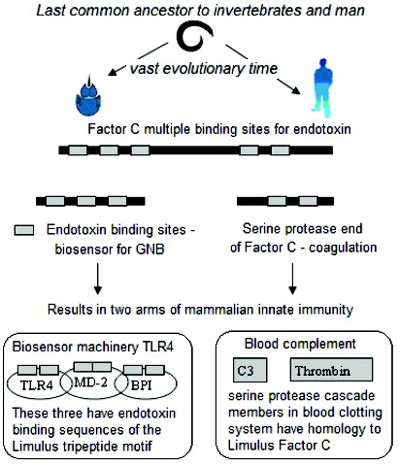 Figure 8. Change occurring over time from invertebrate to mammalian systems results in: (a) conserved endotoxin binding sites ending up in diff erent innate immune system functional groups. (b) more complex mechanisms consisting of more working parts with greater endotoxin specifi city in mammals and more complex activation. (c) a complex anti-coagulation response developed to prevent runaway cascade events.
Figure 8. Change occurring over time from invertebrate to mammalian systems results in: (a) conserved endotoxin binding sites ending up in diff erent innate immune system functional groups. (b) more complex mechanisms consisting of more working parts with greater endotoxin specifi city in mammals and more complex activation. (c) a complex anti-coagulation response developed to prevent runaway cascade events.Another complicating factor in mammalian systems is the existence of symbiotic relationships with large populations of GNB used by the host to various ends. A study by Coats et al [39] looked at the response to pathogen versus symbiont in two closely-related bacterial species.
These two are recognized by mammalian hosts in a counter-intuitive fashion. Bacteroides thetaiotaomicron is a symbiont that outnumbers E. coli 100 to 1 in the flora of the human gut. It metabolizes otherwise indigestible plant sugars. Porphyromonas gingivalis is associated with human periodontal disease. Both synthesize monophosphorilated as well as nonphosphorilated penta-acyl lipid A. The former is a TLR4 weak agonist and the latter is silent altogether. Coats et al. showed that the difference in monophosphorilated lipid A associated with the symbiont versus the pathogen are determined by variant lipid As, 1-phosphorylated versus 4’ phosphorilated penta-acyl lipid A. Surprisingly, it is the pathogen that does not elicit a TLR4 response. This action of avoiding detection allows it to pursue its disease causing activities largely unabated. Similarly, the actions of wellknown plague-associated pathogen Yersina pestis show that the means to evade host defenses can serve pathogenic ends in obtaining a foothold within host defenses. In the fl ea (arthropod) host (27˚C) Y. pestis produces an assortment of lipid A types including hexa-acylated lipid A but in mammalian hosts (37˚C) they produce only tri, tetra, and penta-acyl lipid A with no hexa-acyl lipid A [40]. Therefore, in this case, the “friend” is perceived as an enemy and the “foe” pathogen is received as a friend while harming the host.
Another example of the difficulty of a too-specific biosensor approach to lipid A variable LPS molecules can be seen in a study by Ittig et al [41]. on a particularly potent endotoxin from Capnocytophaga canimorsus that is prevalent in dog bites. They found an unusual case in which the LPS sugar core contributed to TLR4 response. This has seldom been shown to be the case as the core sugar structure is highly conserved. The Ittig et al paper is helpful in summarizing a typical, but by no means uniform, mammalian response as deciphered in recent studies by Park et al [38].
We showed here that C. canimorsus has a penta-acylated lipid A, a feature often correlated to low endotoxicity. In addition, the esterbound 4’ phosphate is lacking. This structural feature is known to reduce the endotoxic activity by a factor of ~100, which can now be better explained based on the recent data obtained with x-ray crystallography on the TLR4/MD-2/LPS complex. In this complex, phosphate groups of lipid A play a crucial role. The 4’ phosphate is thought to bind to positively charged amino acids in the LRR of TLR4 (Arg264, Lys362) as well as to MD-2 (Ser118 and Lys58) in a welldefi ned manner. This ionic interaction seems to be critical for the ligand affinity of lipid A, enabling formation of a hexameric (TLR4/ MD-2/LPS)2 complex necessary for signaling. In the endotoxic lipid A, there is another negatively charged group, 1 phosphate, which binds to positively charged amino acids in the complex, especially in the LRR of both TLR4 and the counter TLR4, called TLR4* (Lys388* of TLR4*, Lys341, Lys362 of TLR4) and also to Arg122 of MD-2. In contrast to the 4’ phosphate which binds to two proteins (TLR4 and MD-2), the 1 phosphate is involved in binding to three proteins in the complex (TLR4, TLR4*, and MD-2), suggesting that this group might be even more important for the formation of a stable hexameric (LPS/TLR4/MD-2)2 complex, as has been reported [41].
Studies comparing chicken (MD-1), horse, mice, and human fine structure of MD-2/TLR4 with man-made amino acid substitutions show exactly how one form can be turned into another (mouse to human etc.) [42, 43]. This level of detail shows just how tedious and apparently arbitrary the host response to LPS can be as a product of evolutionary descent. We take for granted that metazoan’s efforts are an attempt to exclude all lipid A bearing organisms because many GNB are bacterial pathogens.8 However, when we see E. coli and Bacteroides, among the list of prototypical PAMPs included as “foe” while at the very same time recognizing that they are symbiotic guests of mammalian digestive systems, then we see that the host must have a different strategy altogether. In higher organisms, discrimination is occurring in that lipid A is not lipid A “on-off” (presence-absence) as it is as recognized by Limulus but rather lipid A “dialed up or down” with a variety of fine structural settings (TRL4/MD-2 fine detail of acyl, phosphate and sugar structures detected), determined as existing in different tissue types. Raetz [44] called Lipid A an “information-rich molecule” and modern organisms have gained a greater ability to detect the nuances of this complexity than ancient ones. This finely-tuned, differentiated response amounts to a partitioning effect that defines where different organism types are allowed to exist in the body and to what degree, in as much as the host can decide (note that a few organs have “immune privilege” [45]). Included in this localization effort, one can expect to see variants of TLR4, MD-2, and CD-14 fine structure as well. In tissues that do not contain the mechanism (TLR4/MD-2/CD-14 bound or soluble) the response will be of course muted. Munford [46] describes this concept in some depth.
...few of the gram-negative bacteria that have natural habitats in soil and water, or even in the anaerobic microbiome of the GI tract, produce LPS that has hexaacyl LPS. On the other hand, hexaacyl LPS is produced by most of the aerobic or facultatively anaerobic gram-negative bacteria that can inhabit the mucosal surfaces of the respiratory tract and gut. Importantly, the LPSs (lipid As) produced by commensals such as Klebsiella pneumoniae, Enterobacter cloacae, and E. coli are just as “visible” to MD2– TLR4 as are those of mucosal pathogens such as Shigella, Haemophilus, and Neisseria spp. These lipid As have closely similar structures, and they elicit the same inflammatory responses with similar potencies. Clearly, the MD-2– TLR4 receptor does not discriminate between mucosal commensals and pathogens... Here LPS is a virulence factor, since it elicits local inflammatory responses that may damage the host, and yet it also provides a signal that enables the host to prevent further invasion and systemic spread... If our most robust system for sensing aerobic gramnegative bacteria mainly detects ones that can live on our mucosal surfaces, what adaptive role might this have served? One clue is the clinical observation that most mucosal diseases remain mucosal. In other words, mucosal inflammation can be vigorous, and yet systemic dissemination infrequently occurs. The MD-2–TLR4-based response may be disease inducing (in the form of inflammation-associated mucosal damage) and yet protective (preventing bacteremia). It is noteworthy that very little TLR4 has been found in the normal GI epithelium, and it is not expressed on lumenal surfaces except perhaps in colonic crypts. Similarly, MD-2 is minimally expressed by human airway epithelial cells; TLR4 is produced by bladder epithelial cells but not by renal ones. The LPS receptor complex assembles mainly on the defense cells (macrophages, dendritic cells, mast cells, and neutrophils) that patrol the respiratory and gastrointestinal submucosae... Lipid A antagonists might hasten recovery from shigellosis or other mucosal gram-negative diseases, and yet it is also possible that, by diminishing the local inflammatory response, they could favor systemic dissemination.
Summary Discussion
Viewing host strategies from an evolutionary perspective as we have here speaks volumes as to the intention of various biosensor responses to a variety of bacteria that contain a myriad of associated lipid A fine structure types (and many bacteria have more than one type). Since parenteral drugs are invariably introduced via the blood or intrathecally via the spine, then this should inform LAL technologies as to the type of biosensor they should represent. Coming full circle, the prototypical “alarm” of disseminated infection remains E. coli hexaacyl biphosphorilated lipid A. Yet, this alarm system is not simplistic. One need only look at the history of anti-endotoxin therapy candidate drugs to appreciate the complexity of the host-LPS relationship. Eritoran™ is a TLR4 tetra-acylated antagonist developed by Eisai. “Eritoran tetrasodium (E5564) is a synthetic lipopolysaccharide designed to interfere with endotoxin signaling via the TLR4 receptor. The structure of the molecule is based on the lipid A portion of a naturally occurring, weakly agonistic endotoxin found in Rhodobacter sphaeroides” [47]. After several promising studies, Eritoran™ failed its phase III trial (2011). Sepsis treatments have a long history of dashed hopes due to the complexity of the interactions of the immune system with LPS. Companies with promising therapies for sepsis that subsequently experienced dashed hopes stretch back 20 years beginning with Centocor’s Centoxin™, followed by Lilly’s Xigris™, and most recently Eisai’s EritoranTM [48]. Interestingly, each of these drugs has activity against one of the two arms of innate immunity (TLR4 biosensor machinery or Complement system) but none has activity against the effects of endotoxin on both arms.
Here we have seen where the remnants of an ancient endotoxin biosensor has likely ended up in mammalian systems, effectively spread across a wide range of functional parts, and interacting with a multitude of participants that make up the dual system of modern innate immunity. The undifferentiated functionality of an ancient biosensor serves to make the response more general to a broader spectrum of endotoxin types and is therefore less species specific and less subject to downstream exception (as in host machinery polymorphisms). While the prokaryotic cause (LPS) and biosensor detectors (FC and TLR4/MD-2) have remained largely analogous, possibly homologous, the effects in complex host systems have diverged disproportionately. It is the avoidance of complexity via LAL testing that has benefited users and patients the most historically in avoiding confounding, or worse, non-correlated results.
Suffice it to say, LAL users tend to ignore the multitude of accumulating research-oriented data that centers on the modern host response of various species [49]. However, the finer details become relevant when potential new tests seek to increase the specificity of testing to match the level of lipid A fine structure recognized by human immune systems or as needed when trying to validate tests for endotoxin analogs or vaccines9 [50-52]. Increasing biosensor specificity, however necessary it may prove in some instances, is likely to decrease the level of safety in the preclusion of contaminants. Given that not even mouse and human biosensors agree for some of the same endotoxin types, the continued elaboration of species specific responses should not affect the preclusion of contaminants via LAL due to its broader detection spectrum that includes underacylated LPS [39-53]. Stated more directly, evolution has moved biosensor technology forward from the general to the specific, but the specific, as we have seen, also means more idiosyncratic and less inclusive of various contaminant types.
In answering the initial question as to which biosensor technological approach might offer opportunities in testing parenteral drugs, it seems there are a couple of good answers. Firstly, the general approach is necessary to exclude as many endotoxin contaminant types as possible simply because endotoxins that may not elicit host immune responses via activation of TLR4 may still almost certainly bring about other responses (complement activation, coagulation, plasma protein binding, etc.). If one knew which LPS types do not need to be precluded, then the current level of knowledge would be such that sepsis drugs would succeed in their efforts to block all LPS responses. Secondly, a specific approach may find utility where product and process knowledge can inform manufacturers of expected contaminant types that contain atypical LPS structures. Contaminant-type associated LPS may then in some cases be matched with the most relevant test available to preclude them specifically. This later approach is an exciting prospect and one that must be pursued in an informed manner and will almost certainly supplement rather than displace any time soon the broad exclusion of a wide variety of endotoxin types as utilized by LAL technologies.
Author Biography
Kevin Williams, currently at Hospira Inc., has 30 years’ experience in the Pharmaceutical industry specializing in endotoxin testing and control. He has written extensively on the subject of LAL technology including authoring/editing the book “Endotoxins” (Informa Healthcare, 2007). His accomplishments include establishing kinetic BET tests, automation of BET, assay development and validation, depyrogenation validation and troubleshooting. He developed a method of calculating specifications for raw materials based upon establishing proportionality of various constituents in the unit formula of the finished drug product. A previous article in APR, “The BET as a Backdrop for Establishing PAT and RMM Goals”, contrasted the historical development of BET relative to current PAT and RMM efforts.
Acknowledgements
A thank you to Dr. Wyndham W. Lathem of Northwestern University’s Feinberg School of Medicine and Deborah Havlik of Hospira Inc. for reviewing the manuscript.
References
- “Quantitative Comparison of in vitro and in vivo Methods for the Detection of Endotoxin”. J.F. Cooper and J. Levin, Jour. Lab Clin. Med., July 1971, vol.78, No 1, pg. 138-148.
- EndoLISA®: “A Novel and Reliable Method for Endotoxin Detection”. Nature Methods. Hyglos GmbH. Bernried, Germany. Oct. 2011.
- “Assessment of Pyrogenic Contaminations with Validated Human Whole-blood Assay”. Daneshian, von Aulock & Hartung. Nature Protocols 4, 1709 - 1721 (2009).
- “Rational Design of Green Fluorescent Protein Mutants as Biosensor for Bacterial Endotoxin”. Goh, Frecer, Ho and Ding. Protein Engineering,Vol. 16 No. 6., 2002, p. 493-502.
- The Molecular Adsorption-type Endotoxin Detection System Using Immobilized ε -Polylysine, Proc. SPIE 6799, BioMEMS and Nanotechnology III, 679904 (December 27, 2007).
- “Electrochemical Endotoxin Sensors Based on TLR4/MD-2 Complexes Immobilized on Gold Electrodes”. Yeo et al. Biosensors and Bioelectronics. 28 (2011) 139–145.
- “The Molecular Adsorption-type Endotoxin Detection System using Immobilized ε -polylysine”. K. Ooe, et al., Proc. SPIE 6799, BioMEMS and Nanotechnology III. 679904 (December 27, 2007); doi:10.1117/12.759459.
- “Selective Detection of Endotoxin Using an Impedance Aptasensor with Electrochemically Deposited Gold Nanoparticles”. W. Su et al. Innate Immunity. 19 November 2012.
- “Capacitive Microsystems for Biological Sensing”. V. Tsouti et al. Biosensors and Bioelectronics. 27 (2011) 1-11.
- “Endotoxin-Induced Structural Transformations in Liquid Crystalline Droplets”. Lin et al. Science. 2011 June 10; 332(6035): 1297–1300.
- “Structural Prerequisites for Endotoxic Activity in the Limulus Test as Compared to Cytokine Production in Mononuclear Cells”. T. Gutsmann et al. Innate Immunity. 2010 16: 39, June 2009.
- “Lipid A Antagonist, Lipid IVa, is Distinct from Lipid A in Interaction with Toll-like Receptor 4 (TLR4)-MD-2 and Ligand-induced TLR4 Oligomerization”. Saitoh et al. International Immunology. Vol. 16, No. 7, p. 961-969.
- “Including humans in the case of TLR4 polymorphisms, in which they suffer repeat and lifethreatening infections”. Endotoxins. 3rd Ed. Informa Healthcare. P. 102-106
- “Vertebrate Innate Immunity Resembles a Mosaic of Invertebrate Immune Responses”. M. Salzet, TRENDS in Immunology Vol.22 No.6 June 2001.
- “Peeling the Onion: Ribosomes Are Ancient Molecular Fossils”. Hsiao et al. Mol. Biol. Evol. 26(11):2415–2425. 2009.
- “Rescue of Drosophila Labial Null Mutant by the Chicken Ortholog Hoxb-1 Demonstrates that the Function of Hox Genes is Phylogenetically Conserved”. Beat Lutz et al. 176 Genes & Development. 10:176-184. 1996.
- “Demosponge EST Sequencing Reveals a Complex Genetic Toolkit of the Simplest Metazoans”. Matija Harcet. Mol. Biol. Evol. 27(12):2747–2756, 2010.
- “Evidence for the Ancient Origin of the NF-κB/IκB Cascade: Its Archaic Role in Pathogen Infection and Immunity”. Xiao Wei Wang et al. PNAS, Vol. 103 No. 11, 4204–4209.
- “The Oldest Fossil Insect in the World.” Ross, Andrew. Natural History Museum. N.p., n.d. Web. 2013.
- “The Oldest Horseshoe Crab: A New Xiphosurid from Late Ordovician Konservat – Lagerstätten Deposits”. D.M. Rudkin et al. Manitoba, Canada. Palaeontology. Vol. 51, Part 1, p. 1–9. 2008.
- “A Novel Serine Protease Inhibitor Acts as an Immunomodulatory Switch while Maintaining Homeostasis”. N. Jiang et al. J Innate Immun. 2009;1:465–479.
- “The 2.0-Å Crystal Structure of Tachylectin 5Å Provides Evidence for the Common Origin of the Innate Immunity and the Blood Coagulation Systems”. Norman Kairies et al. PNAS. November 20, 2001, Vol. 98 No. 24, 13519–13524.
- “High-affinity LPS Binding Domain(s) in Recombinant Factor C of a Horseshoe Crab Neutralizes LPS-induced Lethality”. N.S. Tan, B. Ho, and J. L. Ding. The FASEB Journal. Vol. 14, 859-870. May 2000.
- “Atomic Force Microscopy Study of the Antimicrobial Action of Sushi Peptides on Gram Negative Bacteria”. Li, Lee, Ho, Ding, and Lim. Biochimica et Biophysica Acta. Vol. 1768, Issue 3, p. 411-418. March 2007.
- “Sushi Peptides: Structural Characterization and Mode of Action Against Gram-negative Bacteria” J.L. Ding, P. Li and B. Ho. Cell. Mol. Life Sci. 65(2008) 1202-1219.
- “Factor C for Treating Gram-negative Bacterial Infection”. Ding, Ho, Tan. Patent US2008008586A1. April 2008.
- “Blood Coagulation: Hemostasis and Thrombin Regulation”. Kenichi A. Tanaka, MD, MSc. et al. Anethesia & Analgesia. Vol. 108, No. 5. May 2009.
- “Plant and Animal Sensors of Conserved Microbial Signatures”. P.C. Ronald and B. Beutler. Science. Vol. 330. 19 Nov. 2010.
- “Interpretation of Biological Activity Data of Bacterial Endotoxins by Simple Molecular Models of Mechanism of Action”. V. Frecer, B. Ho., and J.L. Ding. Eur. J. Biochem. 267, 837-852 (2000).
- “A Structural Perspective on the Interaction between Lipopolysaccharide and Factor C, a Receptor Involved in Recognition of Gram-negative Bacteria”. Takumi Koshiba, Tomoyuki Hashii, and Shunichiro Kawabata, Papers in Press, November 29, 2006.
- “Toll-like Receptors as Targets for Immune Disorders”. Brian Keogh and Andrew E. Parker. Trends in Pharmacological Sciences. July 2011, Vol. 32, No. 7 435.
- “Delineation of Lipopolysaccharide (LPS)-binding Sites on Hemoglobin”. Bahl et al. Jour. Biological Chem. Vol. 286, No. 43, p. 37793–37803, October 28, 2011.
- “The Sole Gateway to Endotoxin Response: How LPS was Identifi ed as TLR4 and its Role in Innate Immunity”. Beutler, B. and A. Poltorak. Drug Metabolism and Disposition, Vol. 29, No. 4 DMD 29:474–478. 2001.
- “Toll to be Paid at the Gateway to the Vessel Wall”. Hansson GK, Edfeldt K. Arterioscler Thromb Vasc Biol. 2005 Jun;25(6):1085-7.
- “Toll to be Paid at the Gateway to the Vessel Wall”. G.K. Hansson, K. Edfeldt, 2005; 25: 1085-1087.
- “Signal Transduction by the Lipopolysaccharide Receptor, Toll-like receptor-4”. E. Palsson- McDermott et al. Department of Biochemistry, Trinity College, Dublin, Ireland - Immunology 2004 113 153–162.
- “Genetic Analysis of Host Resistance: Toll-like Receptor Signaling and Immunity at Large”. B. Beutler et al. Annu. Rev. Immunol. 2006. 24:353-89.
- “The Structural Basis of Lipopolysaccharide Recognition by the TLR4–MD-2 Complex”. Beom Seok Park, Dong Hyun Song, Ho Min Kim, Byong-Seok Choi, Hayyoung Lee, Jie-Oh Lee. Nature. Vol 458: 1191-1195. April 2009.
- “The Lipid A Phosphate Position Determines Differential Host Toll-Like Receptor 4 Responses to Phylogenetically Related Symbiotic and Pathogenic Bacteria”. Coats et al. Infect. Immun., Jan. 2011, Vol. 79, No. 1, p. 203-210.
- “Modification of the Structure and Activity of Lipid A in Yersinia pestis Lipopolysaccharide by Growth Temperature”. Kawahara. Infect. Immun., Aug. 2002, p. 4092–4098.
- The lipopolysaccharide from Capnocytophaga canimorsus reveals an unexpected role of the core-oligosaccharide in MD-2 binding. Ittig S, Lindner B, Stenta M, Manfredi P, Zdorovenko E, Knirel YA, dal Peraro M, Cornelis GR, Zähringer U. PLoS Pathog. 2012;8(5):e1002667. doi: 10.1371/journal.ppat.1002667. Epub 2012 May 3.
- “Crystal Structure of soluble MD-1 and its interaction with lipid Iva”. Yoon et al., PNAS, Vol. 107 No. 24, 2010, p. 10990-10995.º
- “Elucidation of the MD-2/TLR4 Interface Required for Signaling by Lipid”. Iva, Walsh et al. J Immunol. 2008 Jul 15;181(2):1245-54.
- “Bacterial Endotoxins: Extraordinary Lipids that Activate Eucaryotic Signal Transduction”. C. R. Raetz, J Bacteriol. 1993 September; 175(18): 5745–5753.
- “NLRs in Immune Privileged Sites”. Rosenzweig, H., et al. Current Opinion in Pharmacology, Vol. 11, Issue 4, Aug. 2011, pp. 423-428.
- “Sensing Gram-Negative Bacterial Lipopolysaccharides: a Human Disease Determinant?”. Robert S. Munford. Infect. Immun. 2008, 76(2):454.
- “Phase 2 Trial of eritoran tetrasodium (E5564), a Toll-like receptor 4, antagonist, in patients with severe sepsis”. Tidswell et al. Crit. Care Med 2010 Vol. 38, No. 1.
- “The Search for Effective Therapy for Sepsis; Back to the Drawing Board?”. D.C. Angus, MD, MPH, JAMA, December 21, 2011, Vol 306, No. 23.
- “The Evolution of Vertebrate Toll-like Receptors”. J.C. Roach et al. PNAS July 5, 2005 Vol. 102 No. 27.
- “Evaluation of a Whole-Blood Cytokine Release Assay for Use in Measuring Endotoxin Activity of Group B Neisseria meningitidis Vaccines Made from Lipid A Acylation Mutants”. Mark B. Stoddard et al. Clin. Vaccine Immun. Jan. 2010, p. 98–107 Vol. 17, No. 1.
- “Putting Endotoxin to Work for Us: Monophosphoryl Lipid A as a Safe and Effective Vaccine Adjuvant”. Carolyn R. Casella and Thomas C. Mitchell. Cell Mol Life Sci. 2008 October; 65(20): 3231–3240. doi:10.1007/s00018-008-8228-6.
- “Trial Watch FDA-approved Toll-like Receptor Agonists for Cancer Therapy”. E. Vacchelli et al. Oncoimmunology 1:6, 894-907; September 2012; © 2012 Landes Bioscience.
- “Molecular Basis of Reduced Potency of Underacylated Endotoxins”. Teghanemt et al. J. Immunol., 2005, 175:4669-4676.
1The term “biosensor” is often used to describe man-made tools or in vitro methods that utilize natural means of detecting contaminants; however, in this paper I will use the term to include host organism’s mechanisms to protect themselves from gram negative bacterial invaders.
2Hox genes provide a stark example, see [16]
3If this were not true, then the extensive studies distinguishing a wide spectrum of relative reactivity of various natural and synthetic Lipid A fatty acid acyl structures would not make sense.
4Artifacts meaning a residue of a microbial organism and as a contaminant
5Limulus Anti-LPS factor
6Christiane Nüsslein-Volhard’s (Max Planck Institute) reaction to an underdeveloped fl y larvae was “Das war ja toll!” ( “That is weird!”) She coined the name Toll for the mutated gene. The protein product of the Toll gene was necessary for dorsoventral polarity in the fl y (thus the mutation caused deformity) [35].
7Frecer, Ho and Ding: “the LPS-binding sites in proteins may not necessarily be formed by linear amphipathic sequences”. Since these structures loop and wind around then binding sites may not line up to linear sequences.
8Evolution of pathogen and host is a dance, each side changing continuously and attempting to gain a survival advantage.
9Ibid: Today, only three TLR agonists are approved by FDA for use in humans...[52]