Department of Chemical Biology and Therapeutics St. Jude Children’s Research Hospital
Department of Chemical Biology and Therapeutics St. Jude Children’s Research Hospital
Abstract
Carboxylesterases (CE) are ubiquitous enzymes that metabolize ester-containing compounds into the corresponding alcohols and carboxylic acids. This is primarily thought to be a detoxification process for xenobiotics since the proteins are principally localized in cells and tissues likely to be exposed to such agents. However, ester chemotypes significantly improve the water solubility of molecules and hence the strategy of including these functions in poorly soluble drugs is widely employed in the pharmaceutical industry. The problem with such an approach is that these compounds are now subject to the action of CEs, and the hydrolysis effected by these enzymes alters the pharmacokinetics of the agents. Furthermore, recently it has been reported that very potent CE inhibitors are present in natural products, and as a consequence, individuals who take these materials may unwittingly modulate the metabolism and disposition of esterified drugs. In this review, we will discuss the sources of these inhibitors, their chemical structures, their potency towards human CEs and the likely impact they may have on the hydrolysis of small molecules that contain the ester function.
Carboxylesterases
Ester-containing molecules are widely abundant in nature. This functionality can be employed as a structural domain, but it also significantly improves water solubility of small molecules, and consequently this chemotype is present in a very diverse array of chemical structures (see Figure 1). Due to the ubiquitous nature of esterified compounds all organisms, ranging from bacteria to man, have evolved enzymes (carboxylesterases; CE) that can hydrolyze these molecules. This process liberates the corresponding alcohol and carboxylic acid. While the exact purpose for this reaction is unclear, it is though that this represents a detoxification mechanism for xenobiotics1, presumably to allow more facile elimination of the hydrolysis products. To wit, CEs are generally expressed in cells that may be exposed to such agents (e.g. epithelia of the gut and lung, liver, kidney, skin, etc)2,3.
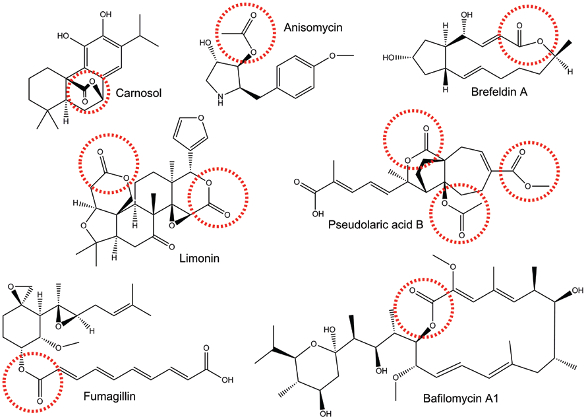 Figure 1. Structures of natural products that contain ester groups (indicated by red dashed circles).
Figure 1. Structures of natural products that contain ester groups (indicated by red dashed circles).In man, three different CEs have been definitively identified (hCE1, hiCE, and hBr3) and the biochemical properties of these proteins have been partially characterized4-6. In general, small substrates (e.g. nitrophenyl acetate) can be hydrolyzed by all three enzymes, whereas larger, more bulky molecules tend to be metabolized by hiCE. The reason for this is largely dependent upon structural constraints enforced by amino acids that form the entrance to the active site of the CEs6. The catalytic amino acids (Ser, His and Glu), that are essential for enzymatic activity, are buried at the bottom of a long, deep, hydrophobic gorge7-9. Hence for substrate hydrolysis, the molecule must enter the ‘mouth’ of the gorge, traverse towards the active site and after reaction, the products must exit the protein. In hiCE, the amino acids surrounding the entrance to the gorge are flexible and hence larger molecules can be accommodated. In contrast, in hCE1, the residues at this location are immobile, and consequently bulky compounds cannot access the catalytic amino acids, resulting in very little, if any, hydrolysis10.
Due to the proclivity of the ester function in natural products, and more recently in synthetically derived drugs (Figure 2), metabolism of these agents by CEs significantly impacts their biodistribution. This activity is further exacerbated by the location of enzyme expression since virtually all routes for drug administration (oral, i.v., topical, etc) results in direct interaction of the small molecules with these proteins. For example, oseltamivir (Tamiflu) is activated by CEs to yield the carboxylate form that acts as an antiviral agent11. Since the parent drug is given orally, activation can occur when passing through the gut epithelia and/or by the enzymes present within the liver during first pass metabolism. Cleary therefore, the biological activity of esterified agents is impacted by the levels, location and the efficiency of enzymatic hydrolysis by CEs.
 Figure 2. The chemical structures of clinically approved drugs that contain ester moieties (indicated by red dashed circles).
Figure 2. The chemical structures of clinically approved drugs that contain ester moieties (indicated by red dashed circles).Inhibition of Carboxylesterases
Ethane-1,2-diones
Until recently, little was known about the selective inhibition of CEs by small molecules. Less specific esterase inhibitors had been identified (typically organophosphorus-based molecules such as pesticides), but they tended to have activity towards a variety of different classes of enzymes (e.g., acetyl and butyrylcholinesterase, lipases, etc). However, following small molecule screening and extensive biochemical and cell biology-based studies, selective CE inhibitors were identified12-17. While several different chemical scaffolds were obtained, the ethane- 1,2-diones were the most potent and tractable, and compounds containing this chemotype have proved to be excellent tools in structural, biochemical and computational experiments.
The prototypical CE inhibitor that contains the ethane-1,2-dione moiety is benzil (Figure 3). This compound demonstrates nanomolar Ki values for human CE inhibition, is inactive towards cholinesterases and is cell permeable14,18. Furthermore, due to the relative simplicity of the molecule, the numerous facile routes for synthesis, and the availability of multiple positions within the structure to allow for analogue generation, reagents generated by such studies have been ideal tools to evaluate CE biology. Using this information, we generated a pharmacophore for CE inhibition and used the derived model to screen libraries of natural products by in silico methods. This approach identified several different classes of abietane diterpinoids (tanshinones and miltirones; Figure 319). Tanshinones are present in large quantities in the Chinese herbal medicine Danshen, a dried root obtained from Salvia miltiorrhiza. The latter is used throughout China, usually drunk as a tea made with hot water, or soaked in Baijiu (a sorghum-based liquor with an alcoholic content of ~50%). Danshen is ‘prescribed’ for many ailments including cardiovascular diseases, ischemia, cirrhosis, glaucoma, bronchitis, wound healing and pancreatitis20.
 Figure 3. The chemical structures of known carboxylesterase inhibitors. The upper row of compounds all contain the ethane-1,2- dione moiety (indicated by the black dashed circles), whereas the lower molecules (fl avanones) lack this functionality.
Figure 3. The chemical structures of known carboxylesterase inhibitors. The upper row of compounds all contain the ethane-1,2- dione moiety (indicated by the black dashed circles), whereas the lower molecules (fl avanones) lack this functionality.Biochemical analysis of organic solvent extracts of Danshen indicated that potent human CE inhibitors were present in this material, and in vitro assays using pure tanshinones confirmed that these molecules could modulate CE activity19. Interestingly, aqueous extraction of these roots failed to yield extracts that contained inhibitors, suggesting that the ‘tea’ described above would be unlikely to affect enzyme activity. However both Baijiu and 50% ethanol can readily extract the tanshinones from Danshen. Since the former is widely used in Asia, individuals may be unwittingly ingesting relatively large amounts of compounds that can modulate the action of human CEs.
CE Inhibitors Lacking the Ethane-1,2-Dione Moiety
Studies by other investigators have identified structurally similar molecules (although lacking the 1,2-dione moiety) and determined that these are CE inhibitors21-23. These typically contain a flavonoid scaffold although it is not entirely clear how they inhibit the enzymes (Figure 4).
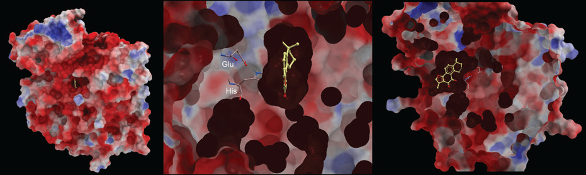 Figure 4. Computational models of tanshinone IIA (yellow) docked into the active site of hCE1. Left panel: A view of the whole protein looking down into the active site gorge. Middle panel: A close up view looking down the gorge. The protein has been sectioned to reveal the glutamic acid and histidine catalytic residues. The serine that initiates the hydrolysis reaction is hidden behind tanshinone IIA. Right panel: A side view of the model displayed in the middle panel.
Figure 4. Computational models of tanshinone IIA (yellow) docked into the active site of hCE1. Left panel: A view of the whole protein looking down into the active site gorge. Middle panel: A close up view looking down the gorge. The protein has been sectioned to reveal the glutamic acid and histidine catalytic residues. The serine that initiates the hydrolysis reaction is hidden behind tanshinone IIA. Right panel: A side view of the model displayed in the middle panel.Structural studies with benzil confirmed that the latter acts as a pseudo-substrate, with the active site serine residues attacking the carbonyl group, but the enzyme cannot break the C-C bond within the dione24. This results in a stabilized tetrahedral intermediate, essentially preventing completion of the hydrolysis cycle. Since similar studies with the flavonoids have not been undertaken, the exact mechanism by which they interact with CE is unknown, but since the active site gorge of the protein is highly hydrophobic, potentially these compounds may localize within this domain, where subsequent hydrogen bonding results in stabilization of the highly oxygenated small molecule. Such a mechanism has been proposed for the inhibition of matrix metalloproteinases and thrombin by sulfonamides [25-28], however determination of the exact interaction of the flavanones and the mode of binding to the CEs will likely require detailed structural studies.
Modulation of CE-Mediated Drug Metabolism by Natural Products
Having determined that organic extracts of Danshen contained potent levels of human CE inhibitors, we postulated that this material may impact esterified drug hydrolysis effected by these enzymes. Indeed this was the case, and the purified tanshinones modulated the antitumor activity of irinotecan, a prodrug that requires CE activation to induce cytotoxicity19. In these in vitro studies, up to 10 times more drug was required to kill tumor cells when treated with tanshinones, as compared to those without. Hence, these diterpinoids can significantly alter the potency of prescribed esterified medications. Since the FDA has recently approved the use of Danshen in clinical trials as part of a formulation called ‘Danshen Dripping Pills’ (12 trials are currently listed for Danshen on www.clinicaltrials.gov), potentially this herbal medicine may modulate the metabolism of other such drugs. Studies to assess the impact of Danshen on CE-mediated drug hydrolysis in vivo are currently underway.
To date, there are no reported studies describing the activities of the flavanones (Figure 3) with respect to esterified drug metabolism. However, if these compounds are cell permeable and have sufficient potency with regard to the relevant substrate, it is likely that they would modulate hydrolysis of these agents.
Potential Natural Product CE Inhibitors
We have continued the in silico search for natural products that contain the ethane-1,2-dione moiety and to date, over 400 such compounds have been identified (see a few selected structures in Figure 5). While the biological activity of these molecules has not been confirmed, it is likely that a significant proportion of them will modulate CE function. The determining factors as to whether they may alter drug metabolism in vivo will be whether such agents are cell permeable, if they are bioavailable, and the potency with which they can inhibit CEs intracellularly. Unfortunately, such studies will likely only be conducted when a drug:drug interaction is observed with a clinically prescribed medicine.
 Figure 5. Natural products containing the ethane-1,2-dione moiety that are likely to be CE inhibitors. The compounds indicated have been identified in marine organisms, fungi, flowering plants and trees.
Figure 5. Natural products containing the ethane-1,2-dione moiety that are likely to be CE inhibitors. The compounds indicated have been identified in marine organisms, fungi, flowering plants and trees.It also should be noted however that biochemical analyses of extracts derived from other Salvia species has confirmed the presence of CE enzyme inhibitors, although the structures of these molecules are currently unknown (Hatfield, Fratt and Potter, unpublished data). Hence, due to the widespread use of natural products/herbal medicines by many individuals and the potential for such material to contain CE inhibitors, we strongly advise against their use in conjunction with esterified drugs.
Summary
The identification of CE inhibitors in natural products is a nascent field, however due to the numerous esterified drugs that are currently used in clinical medicine, and the number of herbal supplements/remedies that are available to patients, it is likely that drug:drug interactions will be identified. To date this has principally concentrated on modulation of drug metabolism by the cytochrome P450s, however since many new clinical entities are being discovered from synthetic chemical libraries, where structural modification with esters is common place, we anticipate that CE-mediated hydrolysis will play an important role in drug pharmacokinetics and disposition. Inhibition of such enzymes will clearly alter these parameters, and hence, we recommend that natural product and herbal supplements be avoided when esterified drugs are used.
Acknowledgments
This work was supported in part by NIH grants CA108775 and AT007531, a Cancer Center Core grant CA21765, and by the American Lebanese Syrian Associated Charities (ALSAC) and St. Jude Children’s Research Hospital (SJCRH).
References
- Cashman J, Perroti B, Berkman C, Lin J. Pharmacokinetics and molecular detoxification. Environ Health Perspect. 1996; 104: 23-40
- Williams FM. Potential for metabolism locally in the skin of dermally absorbed compounds. Hum Exp Toxicol. 2008; 27(4): 277-280
- Hatfield MJ, Tsurkan L, Garrett M, Shaver T, Edwards CC, Hyatt JL, Hicks LD, Potter PM. Organspecific carboxylesterase profiling identifies the small intestine and kidney as major contributors of activation of the anticancer prodrug CPT-11. Biochem Pharmacol. 2011; 81: 24-31
- Humerickhouse R, Lohrbach K, Li L, Bosron W, Dolan M. Characterization of CPT-11 hydrolysis by human liver carboxylesterase isoforms hCE-1 and hCE-2. Cancer Res. 2000; 60: 1189-1192
- Sanghani SP, Quinney SK, Fredenburg TB, Davis WI, Murry DJ, Bosron WF. Hydrolysis of irinotecan and its oxidative metabolites, 7-ethyl-10-[4-N-(5-aminopentanoic acid)- 1-piperidino] carbonyloxycamptothecin and 7-ethyl-10-[4-(1-piperidino)-1-amino]- carbonyloxycamptothecin, by human carboxylesterases CES1A1, CES2, and a newly expressed carboxylesterase isoenzyme, CES3. Drug Metab Dispos. 2004; 32(5): 505-511
- Wadkins RM, Morton CL, Weeks JK, Oliver L, Wierdl M, Danks MK, Potter PM. Structural constraints affect the metabolism of 7-ethyl-10-[4-(1-piperidino)-1-piperidino] carbonyloxycamptothecin (CPT-11) by carboxylesterases. Mol Pharmacol. 2001; 60: 355-362
- Bencharit S, Morton CL, Howard-Williams EL, Danks MK, Potter PM, Redinbo MR. Structural insights into CPT-11 activation by mammalian carboxylesterases. Nat. Struct Biol. 2002; 9: 337-342
- Bencharit S, Morton CL, Hyatt JL, Kuhn P, Danks MK, Potter PM, Redinbo MR. Crystal structure of human carboxylesterase 1 complexed with the Alzheimer’s drug tacrine. From binding promiscuity to selective inhibition. Chem & Biol. 2003; 10: 341-349
- Bencharit S, Morton CL, Xue Y, Potter PM, Redinbo MR. Structural basis of heroin and cocaine metabolism by a promiscuous human drug-processing enzyme. Nat Struct Biol. 2003; 10: 349-356
- Wierdl M, Tsurkan L, Hyatt JL, Edwards CC, Hatfield MJ, Morton CL, Houghton PJ, Danks MK, Redinbo MR, Potter PM. An improved human carboxylesterase for enzyme/prodrug therapy with CPT-11. Cancer Gene Ther. 2008; 15(3): 183-192
- Shi D, Yang J, Yang D, LeCluyse EL, Black C, You L, Akhlaghi F, Yan B. Anti-influenza prodrug oseltamivir is activated by carboxylesterase human carboxylesterase 1, and the activation is inhibited by antiplatelet agent clopidogrel. J Pharmacol Exp Ther. 2006; 319: 1477-1484
- Wadkins RM, Hyatt JL, Yoon KJ, Morton CL, Lee RE, Damodaran K, Beroza P, Danks MK, Potter PM. Discovery of novel selective inhibitors of human intestinal carboxylesterase for the amelioration of irinotecan-induced diarrhea: synthesis, quantitative structure-activity relationship analysis, and biological activity. Mol Pharmacol. 2004; 65(6): 1336-1343
- Wadkins RM, Hyatt JL, Edwards CC, Tsurkan L, Redinbo MR, Wheelock CE, Jones PD, Hammock BD, Potter PM. Analysis of mammalian carboxylesterase inhibition by trifluoromethylketonecontaining compounds Mol. Pharmacol. 2007; 71: 713-723
- Wadkins RM, Hyatt JL, Wei X, Yoon KJ, Wierdl M, Edwards CC, Morton CL, Obenauer JC, Damodaran K, Beroza P, Danks MK, Potter PM. Identification and characterization of novel benzil (diphenylethane-1,2-dione) analogues as inhibitors of mammalian carboxylesterases. J Med Chem. 2005; 48: 2905-2915
- Hyatt JL, Moak T, Hatfield JM, Tsurkan L, Edwards CC, Wierdl M, Danks MK, Wadkins RM, Potter PM. Selective inhibition of carboxylesterases by isatins, indole-2,3-diones. J Med Chem. 2007; 50: 1876-1885
- Hyatt JL, Stacy V, Wadkins RM, Yoon KJ, Wierdl M, Edwards CC, Zeller M, Hunter AD, Danks MK, Crundwell G, Potter PM. Inhibition of carboxylesterases by benzil (diphenylethane-1,2-dione) and heterocyclic analogues is dependent upon the aromaticity of the ring and the flexibility of the dione moiety. J Med Chem. 2005; 48: 5543-5550
- Hyatt JL, Wadkins RM, Tsurkan L, Hicks LD, Hatfield MJ, Edwards CC, Ii CR, Cantalupo SA, Crundwell G, Danks MK, Guy RK, Potter PM. Planarity and constraint of the carbonyl groups in 1,2-diones are determinants for selective inhibition of human carboxylesterase 1. J Med Chem. 2007; 50: 5727-5734
- Hyatt JL, Tsurkan L, Wierdl M, Edwards CC, Danks MK, Potter PM. Intracellular inhibition of carboxylesterases by benzil: Modulation of CPT-11 cytotoxicity. Mol Cancer Ther. 2006; 5: 2281-2288
- Hatfield MJ, Tsurkan LG, Hyatt JL, Edwards CC, Lemoff A, Jeffries C, Yan B, Potter PM. Modulation of Esterified Drug Metabolism by Tanshinones from Salvia miltiorrhiza (“Danshen”). J Nat Prod. 2013; 76(1): 36-44
- Zhou L, Zuo Z, Chow MS. Danshen: an overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J Clin Pharmacol. 2005; 45(12): 1345-1359
- Djeridane A, Brunel JM, Vidal N, YousfiM, Ajandouz EH, Stocker P. Inhibition of porcine liver carboxylesterase by a new flavone glucoside isolated from Deverra scoparia. Chem Biol Interact. 2008; 172(1): 22-26
- Djeridane A, YousfiM, Nadjemi B, Maamri S, Djireb F, Stocker P. Phenolic extracts from various Algerian plants as strong inhibitors of porcine liver carboxylesterase. J Enzyme Inhib Med Chem. 2006; 21(6): 719-726
- Wei Y, Peng AY, Huang J. Inhibition of porcine liver carboxylesterase by phosphorylated flavonoids. Chem Biol Interact. 2013; 204(2): 75-79
- Fleming CD, Bencharit S, Edwards CC, Hyatt JL, Tsurkan L, Bai F, Fraga C, Morton CL, Howard- Williams EL, Potter PM, Redinbo MR. Structural insights into drug processing by human carboxylesterase 1: tamoxifen, mevastatin, and inhibition by benzil. J Mol Biol. 2005; 352: 165-177
- Brandstetter H, Turk D, Hoeffken HW, Grosse D, Sturzebecher J, Martin PD, Edwards BF, Bode W. Refined 2.3 A X-ray crystal structure of bovine thrombin complexes formed with the benzamidine and arginine-based thrombin inhibitors NAPAP, 4-TAPAP and MQPA. A starting point for improving antithrombotics. J Mol Biol. 1992; 226(4): 1085-1099
- MacPherson LJ, Bayburt EK, Capparelli MP, Carroll BJ, Goldstein R, Justice MR, Zhu L, Hu S, Melton RA, Fryer L, Goldberg RL, Doughty JR, Spirito S, Blancuzzi V, Wilson D, O’Byrne EM, Ganu V, Parker DT. Discovery of CGS 27023A, a non-peptidic, potent, and orally active stromelysin inhibitor that blocks cartilage degradation in rabbits. J Med Chem. 1997; 40(16): 2525-2532
- Kiyama R, Tamura Y, Watanabe F, Tsuzuki H, Ohtani M, Yodo M. Homology modeling of gelatinase catalytic domains and docking simulations of novel sulfonamide inhibitors. J Med Chem. 1999; 42(10): 1723-1738
- Whittaker M, Floyd CD, Brown P, Gearing AJH. Design and therapeutic applications of matrix metalloproteinase inhibitors. Chem Rev. 1999; 99: 2735-2776