Protein aggregation is a common deleterious outcome that may occur during the production and storage stages of biotherapeutic protein development. Aggregates are detrimental to drug efficacy, leading to general immunity against the biotherapeutic protein. Protein aggregation is thought to be primarily due to misfolding or partial unfolding, although small protein fragments may also contribute to aggregation initiation. As protein aggregation is a collisional process, increased aggregation is readily observed at high protein formulation concentrations (e.g., 100-240 mg/ml dosages).
Since biotherapeutic proteins are formulated with delivery and storage in mind, it is useful to identify and target regions in the protein structure that are susceptible to localized partial unfolding. These regions may then be addressed by protein engineering strategies designed to stabilize the biomolecule. One potential route towards eliminating large-scale aggregation may involve minimizing nativelike partial unfolding reactions (such as partially unfolded monomers, reversible dimers, and other small-order aggregates) that may be generated under ambient to low temperature storage conditions.
As shown in the above protein stability profile, the region that will most likely generate aggregation-prone species under ambient storage conditions is where rapidly transitioning species are quickly alternating between folded and partially folded states (see Figure 1, green oval). Above Tonset, a greater number of global unfolding reactions may occur. Unfortunately, the reversible transient species that occur within the regions below Tonset are extremely difficult to detect using standard biophysical assays. As a result, there is an impetus to develop methods that enable one to rapidly detect the existence of transient native partially unfolded populations that occur naturally under typical storage temperatures (8 °C to 25 °C) and within physiological temperatures (35 °C to 42 °C).
 Figure 1. Regions of interest for detection of native-like aggregation-prone conformers of therapeutic antibodies. Species that are prominent at the lower temperature range are the folded and partially unfolded forms (In). A greatly increased population of globally unfolded species (Iu) predominate at higher temperatures (above Tonset and at/near the temperature denaturation midpoint, Tm).
Figure 1. Regions of interest for detection of native-like aggregation-prone conformers of therapeutic antibodies. Species that are prominent at the lower temperature range are the folded and partially unfolded forms (In). A greatly increased population of globally unfolded species (Iu) predominate at higher temperatures (above Tonset and at/near the temperature denaturation midpoint, Tm).Current protocols of evaluating and detecting preaggregate states often rely on applying moderate stress conditions to generate relatively stable concentrations of partially unfolded forms for characterization.1,2 These stress conditions include prolonged temperature exposure below Tonset, rapid thermal cycling (e.g., 25 °C to 45 °C, back to 25 °C, repeat), brief pH or low denaturant pulses, and stirring stress. It is presumed that species generated during these moderate stress conditions resemble the authentic “preaggregate” species that drive aggregation reactions during long-term storage. Recently, rapid hydrogen-deuterium exchange quench procedures and advanced mass spectrometry methodologies have emerged as extremely powerful tools for identifying, mapping, and pinpointing the appearance of dynamically fluctuating flexible regions on IgG molecules.2,3 Interestingly, these moderately stressed systems remained in their folded “frustrated” forms (i.e., non-native partially unfolded forms) upon return to ambient temperatures. In most instances, no increases in larger scale aggregation were detected during short (1 hr) moderate temperature stress periods.1,4 The development of these longer lived partially unfolded “frustrated” forms may have some relevance to long-term stability since protein regions that unfold or partially unfold can be detected and identified. If these moderately stressed states are indeed the harbingers of aggregation, it would be necessary to determine if these stressed forms truly resemble the reversible transient states generated at ambient or near physiological temperature conditions.
To detect these reversible transient states, we have employed the use of naturally occurring chaperone proteins. These chaperone proteins are capable of functioning as surveillance proteins due to their ability to recognize, capture, and, in some cases, reverse these trapped partially unfolded states by facilitating unfolding and refolding of the captured protein. Most molecular chaperones are responsible for preventing large-scale aggregation under physiological temperature and solution conditions.
One of the most promiscuous chaperones that binds either partial or mostly unfolded forms is the Group I (i.e., of eubacteria/mitochondrial origin) chaperonin protein GroEL, which is isolated from E. coli. The nucleotide-free form of GroEL binds to partially folded proteins with high affinity where the binding constant may be determined by the extent of hydrophobic surface exposed on the substrate protein. This is due to the ability of the chaperonin to bind to partially folded protein forms through its multiple interaction sites within its large 45 Å hydrophobic binding pocket (Figure 2).
We have found that the chaperonin protein GroEL can bind and kinetically capture transient partially unfolded states that occur within ambient physiological temperature ranges. In Figure 2, we illustrate a chaperonin protein (GroEL from E. coli) binding to a partially folded antibody protein (in this case through its Fc portion). The strength of the dissociation binding constants (Kd ) for such interactions can approach those observed in antibody-antigen interactions. In vivo, these protein-substrate interactions are naturally reversed by nucleotide binding (ATP) along with a co-chaperonin species. Our laboratory has repeatedly shown that transient partially unfolded forms of a substrate protein may freely bind through in-solution diffusion to an immobilized form of the chaperonin.1, 4-6 We weaken protein-substrate rebinding and resume refolding of the substrate protein by adding ATP or an ATP/osmolyte solution to the chaperoninsubstrate system.7 Our initial studies have demonstrated that one may detect the presence of dynamic transient preaggregates from simply incubating moderately stressed mAb proteins with GroEL in solution, in some cases forming GroEL-mAb complexes can be readily observed using negative stain electron microscopy1,4
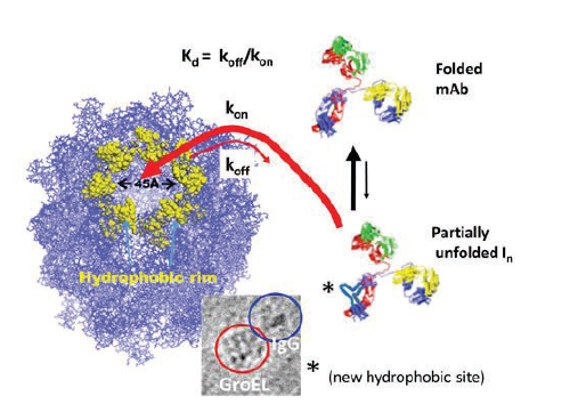 Figure 2. Depiction of a chaperonin (GroEL)-antibody (IgG) binding reaction. The chaperonin recognizes transient or stable hydrophobic patches that occur as a protein substrate undergoes partial unfolding. This binding can occur even in situations where partial unfolding reactions are transient. With binding affi nities and association rates approaching diff usion rates, (due to the large 45 Å chaperonin binding surface), the chaperonin can essentially trap these transient hydrophobic states as they fl icker into existence. The electron microscopy inset shows a side view of a GroEL molecule binding to the Fc portion of Human IgG molecule (Fab domains remain visible).
Figure 2. Depiction of a chaperonin (GroEL)-antibody (IgG) binding reaction. The chaperonin recognizes transient or stable hydrophobic patches that occur as a protein substrate undergoes partial unfolding. This binding can occur even in situations where partial unfolding reactions are transient. With binding affi nities and association rates approaching diff usion rates, (due to the large 45 Å chaperonin binding surface), the chaperonin can essentially trap these transient hydrophobic states as they fl icker into existence. The electron microscopy inset shows a side view of a GroEL molecule binding to the Fc portion of Human IgG molecule (Fab domains remain visible).In order to automate the detection of transient partially folded states as they dynamically fluctuate in solution, we have developed a GroEL biosensor system specifically designed with bio-layer interferometry (BLI) instrumentation to capture and detect the presence of these transient forms at ambient or near physiological temperatures (Figure 3)1,4 In processes that are equivalent to in-solution capture (Figure 2), the GroEL-BLI biosensor system detects and captures these dynamic, transient partially unfolded mAb forms by measuring a change in binding amplitude due to GroEL-mAb interactions (Figure 3). The easy reversal of the captured IgG tightly bound to the chaperonin biosensor by an addition of ATP or an ATP/osmolyte solution affirms that the binding reaction is specific.
 Figure 3. The GroEL biosensor binds protein forms that transiently expose hydrophobic patches along the protein surface. The output of preaggegate or aggregation-prone IgG binding is shown on the right as a BLI sensorgram. The measurable parameters from this output can yield association rates (kon) and dissociation rates (koff ). The ratio of koff /kon yields an apparent dissociation constant (Kd). Specifi c binding is verifi ed by the ability to reverse binding by adding ATP or an ATP/osmolyte mixture to the solution. The amplitude of the fi nal value of the on rate kinetic trace (green circle) is directly proportional quantitatively to the amount of protein with hydrophobic patches.
Figure 3. The GroEL biosensor binds protein forms that transiently expose hydrophobic patches along the protein surface. The output of preaggegate or aggregation-prone IgG binding is shown on the right as a BLI sensorgram. The measurable parameters from this output can yield association rates (kon) and dissociation rates (koff ). The ratio of koff /kon yields an apparent dissociation constant (Kd). Specifi c binding is verifi ed by the ability to reverse binding by adding ATP or an ATP/osmolyte mixture to the solution. The amplitude of the fi nal value of the on rate kinetic trace (green circle) is directly proportional quantitatively to the amount of protein with hydrophobic patches.The data from the output of every individual biosensor sensorgram yields the association/dissociation kinetics (i.e., formation and release of initial GroEL-mAb collision complexes) and, more importantly, apparent Kd estimates (koff /kon) for binding interactions. This latter metric enables one to determine if the preaggregation transient states detected by the GroEL biosensor at ambient temperatures are comparable to the longer-lived partially unfolded states generated under moderate stress conditions (amplitude match adjusted). In simpler terms, similar partially unfolded states may lead to similar Kd values since these Kd values are determined primarily by the amount of exposed hydrophobic regions that occur as particular mAb regions partially unfold. The GroEL biosensor output is also useful in assessing the relative amount of partially folded dynamic transients in solution as functions of both concentration and temperature. In addition to the information within one BLI sensorgram, other global thermodynamic parameters such as activation energies and simple Van ’t Hoff relationships (assuming heat capacity changes are minimal within low temperature ranges) can be obtained by evaluating GroEL biosensor kinetic partitioning data within small non-denaturing temperature ranges (e.g., 15 °C to 45 °C).
To date, the ability to detect transient states at ambient or near physiological conditions has been particularly useful for detecting the existence of preaggregate states of mAb and bispecific mAb constructs before any large-scale aggregation is even observed.1,4 Furthermore, the interactions between GroEL and mAbs either in solution or released from the biosensor are easily visualized using negative stain electron microscopy. One potentially exciting outcome of GroEL biosensor detection and EM visualization is that, in some instances, the flexible regions pinpointed by hydrogen-deuterium exchange mass spectrometry (H/D Exchange-MS) correlate with the same regions that interact with the chaperonin.1,2
Evaluating Kinetic Stabilities of Aggregation-Prone Proteins with Chaperonin-BLI Denaturant Pulse
The kinetic stabilities of aggregation-prone proteins or wild type and mutant protein comparisons may also be evaluated using the chaperonin bio-layer interferometry technology.8,9 In this methodology, the attachment scheme is reversed such that the target protein of interest is attached to the biosensor surface via covalent or metal chelation methods in an orientation-specific manner. The immobilized protein is then subjected to a short exposure to a chemical denaturant ladder using the dip and read technology of the bio-layer interferometry system. The denaturant is promptly washed away in a 10 second wash step and the amount of partially or completely unfolded protein is immediately evaluated by detecting the amount of chaperonin (GroEL) that binds to the affected protein population (Figure 4A). The GroEL binding amplitude is plotted as a function of increasing denaturant concentration in order to generate a kinetically controlled denaturation isotherm. Comparative kinetic denaturation isotherms of a native protein and a missense variant maltose binding protein (MBP) are shown in Figure 4B.
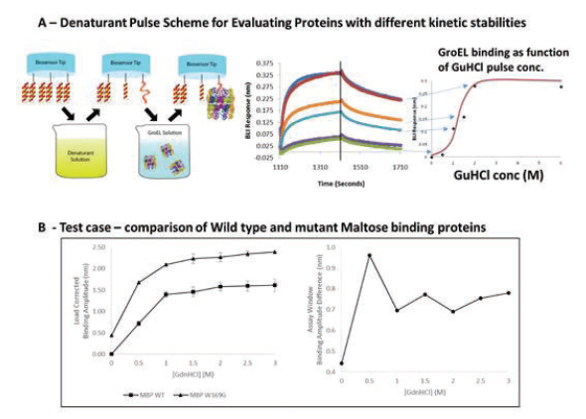 Figure 4. A) Denaturant pulse scheme. BLI biosensors containing the target protein are dipped into a denaturant for a defi ned interval of time, washed to remove any residue denaturant, and then dipped into a chaperonin solution where binding reactions to partially unfolded proteins may occur. The chaperonin binding amplitudes are recorded and plotted as a function of denaturant concentration to yield a kinetic denaturation isotherm (far right). B) Comparative kinetic denaturation isotherms in triplicate of a wild type maltose binding protein with a W169G missense variant shows that the wild type protein is more kinetically stable than the mutant protein. Experiments were performed using the Pall FortéBio Octet RED96E biolayer interferometry (BLI) system.
Figure 4. A) Denaturant pulse scheme. BLI biosensors containing the target protein are dipped into a denaturant for a defi ned interval of time, washed to remove any residue denaturant, and then dipped into a chaperonin solution where binding reactions to partially unfolded proteins may occur. The chaperonin binding amplitudes are recorded and plotted as a function of denaturant concentration to yield a kinetic denaturation isotherm (far right). B) Comparative kinetic denaturation isotherms in triplicate of a wild type maltose binding protein with a W169G missense variant shows that the wild type protein is more kinetically stable than the mutant protein. Experiments were performed using the Pall FortéBio Octet RED96E biolayer interferometry (BLI) system. Opportunities for Evaluating Preaggregate, Transiently Destabilized States with GroEL Biosensors
As we expand the ability to assess preaggregate and transient partially unfolded protein forms, determining whether strong correlations exist between flexible mAb sites identified by H/D Exchange-MS and GroEL-BLI biosensor binding responses will become necessary. In particular, it would be extremely useful to determine if measured apparent binding constants (Kd s) to the GroEL biosensor reflect the extent of hydrophobic surface changes that are assessed by H/D Exchange-MS mapping. For instance, one could determine if the binding affinities directly correlate with increases or decreases in regions of local flexibility with respect to denaturing or stabilizing excipient solutions. Finally, changes in moderately stressed mAb binding to the GroEL biosensors with time may reflect reversible but slow hysteresis effects in perturbed states. This is possible since the GroEL biosensor technology is able to detect diminished binding due to slow reversibility of perturbed states to their native folded forms in the absence of any measurable aggregation. With the ability to demonstrate linearity with respect to concentrations and assess apparent binding constants, it will also be possible to determine if moderately stressed states are equivalent or different to the transient states that partition onto the GroEL chaperonin under ambient temperature conditions.
References
- Pace SE, Joshi SB, Esfandiary R, Stadelman R, Bishop SM, Middaugh CR, Fisher MT*, Volkin DB*. (2018). The Use of a GroEL-BLI Biosensor to Rapidly Assess Preaggregate Populations for Antibody Solutions Exhibiting Diff erent Stability Profiles. J Pharm Sci. 107(2):559-570.
- Toth RT 4th, Pace SE, Mills BJ, Joshi SB, Esfandiary R, Middaugh CR, Weis DD, Volkin DB. (2017) Evaluation of Hydrogen Exchange Mass Spectrometry as a StabilityIndicating Method for Formulation Excipient Screening for an IgG4 Monoclonal Antibody. J Pharm Sci. Dec 18. [Epub ahead of print].
- Majumdar R, Esfandiary R, Bishop SM, Samra HS, Middaugh CR, Volkin DB, Weis DD. (2015) Correlations between changes in conformational dynamics and physical stability in a mutant IgG1 mAb engineered for extended serum half-life.MAbs. 7(1):84-95.
- Naik S., Kumru OS., Cullom M., Telikepalli SN., Lindboe E., Roop TL., Joshi SB., Amin D., Gao P., Middaugh CR., Volkin DB, and Fisher MT (2014) Probing structurally altered and aggregated states of therapeutically relevant proteins using GroEL coupled to Bio-Layer Interferometry, Protein Science 23:1461-78.
- Correia, AR., Naik, S., Fisher, MT., and Gomes, CM., (2014) Probing the Kinetic Stabilities of Friedreich’s Ataxia Clinical Variants Using a Solid Phase GroEL Chaperonin Capture Platform Biomolecules 2014, 4(4), 956-979.
- Smith K.E., Voziyan P.A., and Fisher M.T. (1998) “Partitioning of rhodanese onto GroEL: Chaperonin binds a reversibly oxidized form derived from the native protein” The Journal of Biological Chemistry 273 28677-28681.
- Voziyan P.A. Jadhav, L and Fisher M.T. (2000) “Refolding a glutamine synthetase truncation mutant in vitro: identifying superior conditions using a combination of chaperonins and osmolytes.” The Journal of Pharmaceutical Sciences 89, 1036-1045.
- A Lea, WA, Naik, S., Chaudhuri, T., Machen, AJ., O’Neil, PT, McGinn-Straub, W Tischer, A., Auton, MT., Burns, JR., Baldwin, MT., Khar, KR., Karanicolas J and Fisher MT. (2016) Chaperonin-based biolayer interferometry to assess general the kinetic stability of metastable, aggregation-prone proteins. Biochemistry, 2016, 55 (35), 4885–4908.
- O’Neil, PT., Machen, AJ., Thompson, JA., Wang, W, Hoang, QQ., Baldwin, MT., Khar, KR., Karanicolas J., and Fisher MT., (2018) “Constructing Kinetically Controlled Denaturation Isotherms of Folded Proteins Using Denaturant-Pulse Chaperonin Binding” Methods in Molecular Biology Protocols in Press.