Introduction
The monoclonal antibodies (mAbs) market has changed rapidly in the past five years, with mAbs representing more than 50% of the overall biotherapeutic market. Now, speed to clinic is a huge determinant of success, much of the mAb industry has migrated to platform approaches for discovery, product and process development, and mAbs therapeutics have demonstrated themselves to be particularly well suited toward the application of platform approaches. The platform providers help to optimize the product and process development that keeps improving the production yield and efficiency, and also streamlines the regulatory and quality records that allow industry and regulators alike to move faster and with greater consistency. As of December 2019, 79 therapeutic mAbs have been approved by the FDA, and approximately 25 mAbs are under review by FDA and EMA; among them, ~60% of these mAbs belong to IgG1 class.
Despite such progress, instability associated with various critical quality attributes continues to be a major concern for the mAbs industry, as it can either directly or indirectly affect mAbs biological activity and efficacy. Risks to protein stability can arise from not only a low percentage of ‘foreignness’ of the protein, but also from the formation of aggregates and degradants during manufacture and long-term storage conditions, where proteins are exposed to physical and chemical stress conditions. The most common causes of physical degradation encountered during manufacturing, processing and ultimately clinical administration include excessive temperature, mechanical, and freeze/ thaw stresses. Chemical treatments (chemical modifications, pH, and light exposure etc.) induced degradation affect the primary sequence and may lead to significant changes in structure. As a result, forced degradation studies have been commonly used by the industry to support the development of mAb therapeutics throughout the life-cycle of the products. It is also the expectation of agencies that forced degradation studies be used to understand the product degradation pathways, and establish stability indicating methods enabling monitoring degradation, if it occurs, within the shelf life. The information gathered can provide highly relevant data for our better understanding of the biochemical and biophysical nature of the molecules to ensure that products are well characterized in a timescale that enables rapid clinical and commercial development. A thorough understanding of the mechanisms for the causes of mAb degradation could allow crafting stabilization strategies, and facilitate the development of efforts to improve the stability.
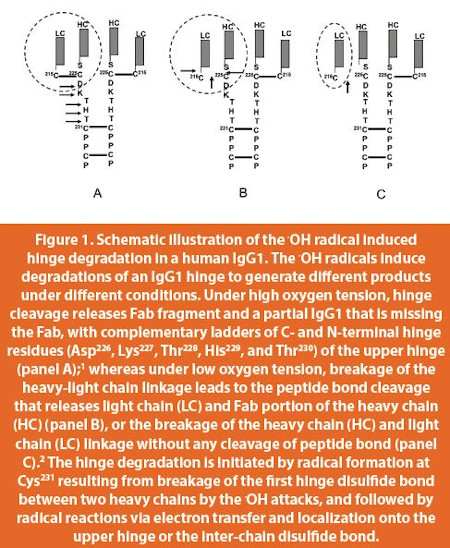
This review focuses on hydroxyl radical induced degradation that occurs in the hinge region of an IgG1, resulting in a variety of products under different reaction conditions.1,2 The hinge region refers to the sequences from the core hinge with a sequence of Cys231-Pro-Pro-Cys through the upper hinge with a sequence of Asp226-Lys-Thr-His-Thr within the heavy chains, to the disulfide bond between Cys 225 of the heavy chain and Cys 215 of the light chain that connect the heavy and light chains, as shown in Figure 1. As demonstrated by Garrison3 Davies and Dean4 that radical reactions can take different pathways under different oxygen tensions, that can be related to different physical and chemical conditions. Under high oxygen tension, the hinge cleavage releases degraded products consisting of a Fab domain and a partial IgG1 that is missing the Fab,1 whereas under low oxygen tension, products are generated by either cleavage of the peptide bond between the light chain (LC) and the heavy chain (HC), or just releasing a LC without any cleavage of peptide bond2 (Figure 1).
This is a radical driven mechanism that specifically attacks Cys231 first in the core hinge sequence of Cys231-Pro-Pro-Cys of an IgG1, leading to the formation of various degraded products under different conditions. Since all four classes of IgG molecules share a very similar hinge sequence (Table 1), therefore, this mechanism not only works for all IgG1 mAbs, but also potentially works for other classes of mAb molecules as well. In addition, the radical induced degradation mechanism could explain many degradation products generated under physiochemical conditions, therefore, it provides new insights into how to engineer a new generation of therapeutic IgG1 that is capable of resisting the radical related degradation from manufacture and stability perspective.
Hydroxyl Radical and H2O2
As precursor of hydroxyl radical (.OH), H2O2 involves in a lot of the trace reactions in vivo that can occur in cell-based cell culture systems, and there is convincing evidence linking them to the H2O2 generation. Most recombinant mAbs are produced by Chinese hamster ovary (CHO) cells using the bioreactor cell culture system that is the industrial standard platform approach, where the conditions mimic the physiological environment, providing in vivo-like conditions. It is well established that mitochondria are the main source of reactive oxygen species generation in cells).5,6 The rate of H2O2 production in isolated mitochondria is 0.6–1.0 nmol/mg min.6 In addition, it has been recently demonstrated that antibodies have the ability to catalyze the generation of H2O2 in a highly efficient manner.7,8 These conditions are sufficient to be the source of H2O2, creating a high oxygen tension condition with a local concentration high enough for hydroxyl radical (.OH) formation around mAb molecules that are produced by CHO cells. A similar low oxygen tension exists, where H2O2 is a derivative product of superoxide radical (O2-..) under thermal incubation conditions. The O2-.. is the first intermediate product in a chain reaction of the reduction of molecular oxygen to water,9 and it can be converted to H2O2 by redox active metal ions and/or superoxide dismutase (SOD).2 Estimated in vivo concentrations of H2O2 and O2-.. have been reported as 10-7 to 10-9 M and 10-11 M, respectively,5 whereas these two species do not interact to produce hydroxyl radical.10,11 In the chain reaction of the reduction of oxygen into water, the overall reaction rate is proportional to collision frequency, so O2-.. and H2O2 fluxes depend directly upon the ambient concentration of oxygen.9 Thus, low oxygen tension may allow thermal incubation induced radical reactions to take place via diff erent routes and generate products diff erent from those generated under high oxygen tension.
Because trace amounts of redox metals (e.g. Fe 2+/3+ and Cu+/2+) are always present in solvents or may be bound to proteins, their concentration could be high enough to function as catalysts for the generation of radicals. This hypothesis is consistent with previous observations that redox metal ions play an important role in site-selective radical attack either by binding to a protein or staying free in solution. In both cases, the redox metal ions accelerate the reaction by catalyzing the generation of hydroxyl radicals through the Fenton chemistry reactions, as demonstrated by our experimental results.1,2
Radical Formation and Transfer on Hinge Residues
The radical induced degradation can be initiated by the formation of a OH. via a reaction of a redox metal ion with H2O2. In the next step of the reaction, the OH. specifically attacks the core hinge of Cys231-Pro-Pro-Cys by breaking the first interchain disulfide bond between Cys231 residues of two heavy chains, yielding radical formation in one of the Cys231 side chains (thiyl radical), which functions as the primary radical center, and oxidation to sulfonic acid in the other. With the radical formation at Cys231, the electron transfer (ET) will occur to release an elevated energy, and it requires hydrogen bond network which can be formed by side chains among amino acids in the hinge. The upper hinge with a sequence of Asp226-Lys-Thr-His-Thr is much preferred for the radical localization than the core hinge with a sequence of Cys231-Pro-Pro-Cys, due to the fact that Pro does not have a side chain.1
His229 residue in the upper hinge plays a critical role in the radical induced degradation, the imidazole ring of His not only participates in the hydrogen bond that maintains the stability of the upper hinge region, but also facilitates the radical reaction after localizing the electron transferred from the Cys231 radical. The imidazole ring forms a new transient radical center that is capable of transferring the electron by extracting a proton from its neighboring residues, leading to hinge cleavage. A typical electron transfer (ET), an energy-driven process, usually does not result in the cleavage of a peptide bond. Nonetheless the ET in the hinge is a localized event because the upper hinge is framed by disulfide bonds at its N and C termini and lacks structurally proximal residues capable of forming a hydrogen bond network. The disulfide bonds that frame the upper hinge block the release of the elevated energy at a new radical site. Consequently, the hinge cleavage is the outcome of an energy-driven process in the hinge region upon the radical attack.
Subscribe to our e-Newsletters
Stay up to date with the latest news, articles, and events. Plus, get special offers
from American Pharmaceutical Review – all delivered right to your inbox! Sign up now!
Under thermal incubation, or a low oxygen tension condition, an electron transferred from Cys231 may directly localize onto the disulfide bond between Cys 215 of the light chain (Cys 215 L) and Cys225 of the heavy chain (Cys 225 H) that connects the heavy and light chains (HC-LC bond, Figure 1). As indicated by Davies and Delsignore,12 radical attacks results in gross distortions of secondary and tertiary structures of a protein, our results confirmed the conclusion and suggested that radical reactions under the thermal incubation conditions also alter local conformation or conformational dynamics of an IgG1. The new local conformation or conformational dynamic in the upper hinge region could bring the H-L bond more proximal to Cys231, which would allow interaction between the electron and the H-L bond.2 Although the distance between Cys231 and the HC-LC bond of 16–17Å based on structural model is greater than a typical electron tunneling distance of 14 Å,13 the extreme flexibility of the upper hinge could allow a direct transfer of electron that brings the HC-LC bond more proximal to Cys231 to facilitate the radical reactions.2

Radical Mechanism of Hinge Degradation
It remains to be determined which of the two hinge Cys231 residues in the heavy chains forms the radical, whereas once it is formed, the electron transfers, localizes at His229, and forms a new radical center that is capable of picking up a proton from its neighboring residues. The high degree of flexibility of the upper hinge allows greater conformational dynamics than in the rest of the IgG1 molecule and facilitates the formation of the hydrogen bond involving the imidazole ring. The imidazole ring forms a new transient radical center that is capable of transferring the electron by extracting a proton from its neighboring residues, leading to hinge cleavage at a one-radical cleavage per molecule basis to release the Fab domain. The fact that only one of the two chains of the hinge was cleaved suggests the coupling of one thiyl radical with one imidazole ring, and the chain being cleaved would be determined by the thiyl radical on the same chain.
Different from a typical peptide bond hydrolysis that breaks a CO-NH bond, radical induced cleavage occurs at α-carbon that is next to the CO-NH bond, protein backbone fragmentation under high oxygen tension can be initiated via two major pathways: the diamide and α-amidation pathways. In the diamide pathway, the cleavage at the α-carbon of the peptide bond yields an isocyanate derivative at the heavy chain N terminus of the C-terminal (Fc-HC) and a ketone carbonyl function at the heavy chain C terminus of the N-terminal backbone fragment. The α-amidation pathway is characterized by the formation of an N-terminal α-ketoacyl derivative at the C-terminal backbone fragment (Fc-HC) and a diamide function at the C terminus of the N-terminal backbone fragment (Scheme 1). These degraded products with adducts at new C- or N-termini can be detected by mass spectrometry, therefore, present the experimental evidence of the radical mechanisms for the hinge degradation of an IgG1.1

On the other hand, under low oxygen tension, electron addition to the H-L bond results in the formation of a disulfide bond radical anion. Decomposition of the anion results in breakage of the C–S bond of the side chain of Cys225HC or Cys215LC that is in the H-L bond, as a result, some sulfur groups are oxidized, and the rest remain in a reduced form. In the counterpart, the Cys that lost the sulfur group forms a carbon-centered radical that is the same species as a result of hydrogen abstraction from the side chain of Ala, forming unstable dehydroalanine as described in Scheme 2. Hydrolysis of the dehydropeptide yields an amide at the N-1 position of the new C terminus.2 Subsequent radical reactions and hydrolysis reactions lead to cleavage of the peptide bond, yielding an amide at the new C terminus. Such reactions are slow, with low yields of minor products arising from cleavage of the C-S-C link, but produce significant amounts of cleavage products under thermal incubation conditions.2

Implication for Stability and Other Classes of IgG Molecules
With the accumulating knowledge and experience with IgG mAb therapeutics, more effort has been focused on exploring and extending the variety of antibody structures that can improve product quality and efficacy to better serve patients, like ADC, bispecific and Fc fusion therapeutics. However, the application of antibody engineering strategies to all mAbs tends to be limited, as structure or modifications may be optimized for one mAb, but may compromise another in vivo. Thus, a mechanism-based strategy for engineering mAbs to improve multiple properties and/or functions should be more successful in delivering the development and manufacturing goals.
Our results indicate that it is feasible to design a new IgG1 with a hinge that is more resistant to radical induced degradation while maintaining its biological activity. The substitution of the hinge His229 residue with a polar residue (Ser and Asn) blocked the radical-mediated hinge cleavage.14 The variants were more stable than the native molecule and maintain the in vitro biological functions of the IgG1 (e.g. FcγR binding and caspase-3 assay).15 In addition, substitution of His with Tyr reveals a potential strategy for improving stability and function of the IgG1: it maintains a similar pharmacokinetic profile with respect to the native molecule and increases the antibody binding to FcγRIII receptors by 2–3 fold, and improves ADCC activity by 2-fold,15 demonstrating the feasibility of engineering the upper hinge to improve the stability and effector function of the IgG1.
There is a highly conserved core sequence of the hinge CPXC between IgG molecules as shown in Table 1, implying that other classes of antibodies also could undergo similar radical induced degradation reactions, particularly for the IgG3 subtype that has an upper hinge sequence similar to IgG1; however, a much longer length of the upper hinge of IgG3 (double length than IgG1) may reduce the frequency of a radical / electron interacting directly with the H-L bond due to a much longer distance for the electron transfer. For an IgG4, a sequence of Pro-Pro proceeding to the core hinge of Cys-Pro-Ser-Cys could prevent an ET to the upper hinge, consequently, resulting in formation of a half molecule upon radical attack. Based on such knowledge, designing a new generation of IgG1 antibodies with a new upper hinge sequence that is more resistant to radical attack could not only improve the yield of production but also potentially enhance the stability and efficacy profile in vivo.
References
- Yan B, Yates Z, Balland A, and Kleemann G R. Human IgG1 hinge fragmentation as the result of H2O2-mediated radical cleavage. J. Biol. Chem. 2009, 284: 35390–35402.
- Yan B, and Boyd D. Breaking the light and heavy chain linkage of human immunoglobulin G1 (IgG1) by radical reactions. J. Biol. Chem. (2011) 286: 24674–24684
- Garrison W M. Reaction mechanisms in the radiolysis of peptides, polypeptides, and proteins. Chem. Rev (1987) 87, 381–388
- Davies MJ, and Dean R T Radical-mediated Protein Oxidation (1997) Oxford University Press, Oxford, U.K. pp. 50–120,
- Chance B., Sies H., and Boveris A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev.(1979) 59: 527–605
- Boveris A., and Chance B. The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem. J. (1973) 134: 707–716
- Zhu X, Wentworth P Jr, Wentworth AD etc. Probing the antibody-catalyzed water-oxidation pathway at atomic resolution Proc. Natl. Acad. Sci. U.S.A. (2004) 101: 2247–2252
- Wang PX., and Sanders PW. Immunoglobulin Light Chains Generate Hydrogen Peroxide J. Am. Soc. Nephrol. (2007) 18, 1239–1245
- Imlay J A. Cellular defenses against superoxide and hydrogen peroxide. Annu. Rev. Biochem. (2008) 77, 755–776
- Rosen GM, Pou S, Ramos CL, Cohen MS, and Britigan BE. Free radicals and phagocytic cells FASEB J. (1995) 9: 200–209
- MacManus-Spencer LA, and McNeill K. Quantification of singlet oxygen production in the reaction of superoxide with hydrogen peroxide using a selective chemiluminescent probe J. Am. Chem. Soc. (2005) 127: 8954–8955
- Davies KJ, Delsignore ME., and Lin S W. Protein damage and degradation by oxygen radicals J. Biol. Chem. (1987) 262: 9902–9907
- Page CC, Moser CC, Chen X, and Dutton P L. Natural engineering principles of electron tunneling in biological oxidation-reduction. Nature, (1999) 402: 47–52
- Yates Z, Gunasekaran K, Zhou H, Hu Z, Liu Z, Ketchem RR, and Yan B. Histidine residue mediates radical-induced hinge cleavage of human IgG1. J. Biol. Chem. (2010) 285, 18662–18671
- Yan B, Boyd D, Kaschak T, Engineering Upper Hinge Improves Stability and Effector Function of a Human IgG1. J. Biol.Chem. (2012), 287: 5891–5897,
Author Biography
Bryant Yan, corresponding author, is a Sr. Director at Acceleron Pharma, in charge of analytical development and quality control within technical operation. Byan@acceleronpharma.com, 1-617 649-9237, www.acceleronpharma.com.